説明文の1
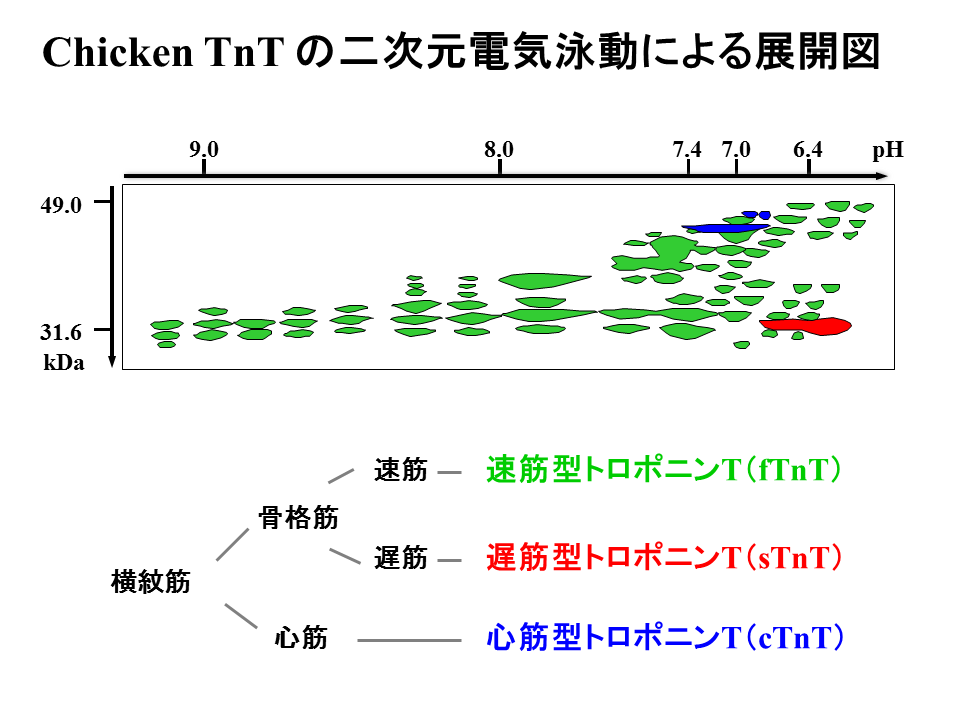
トロポニンT遺伝子は選択的エクソンをもち、そのスプライシングや翻訳後の修飾(リン酸化)により、多くのアイソフォームを産生する。それらのアイソフォームは二次元電気泳動法により、等電点約6.0から9.5、分子量約30から50kDa(ダルトン)の領域に存在する。
説明文の2
トロポニンTは、速筋型トロポニンT(fTnT)、遅筋型トロポニンT(sTnT)、心筋型トロポニンT(cTnT)の3種類から生み出される。速筋型遺伝子は27個のエクソンをもち、そのうち16個が選択的スプライシングを受ける可能性のあるエクソンである。それゆえ、速筋型遺伝子から27すなわち128とおりのパターンのアイソフォームが生み出される可能性がある。遅型型遺伝子は14個のエクソンをもち、心筋型遺伝子は18個のエクソンをもつが、両者とも選択的スプライシングを受ける可能性のあるエクソンは1個だけである。
説明文の3
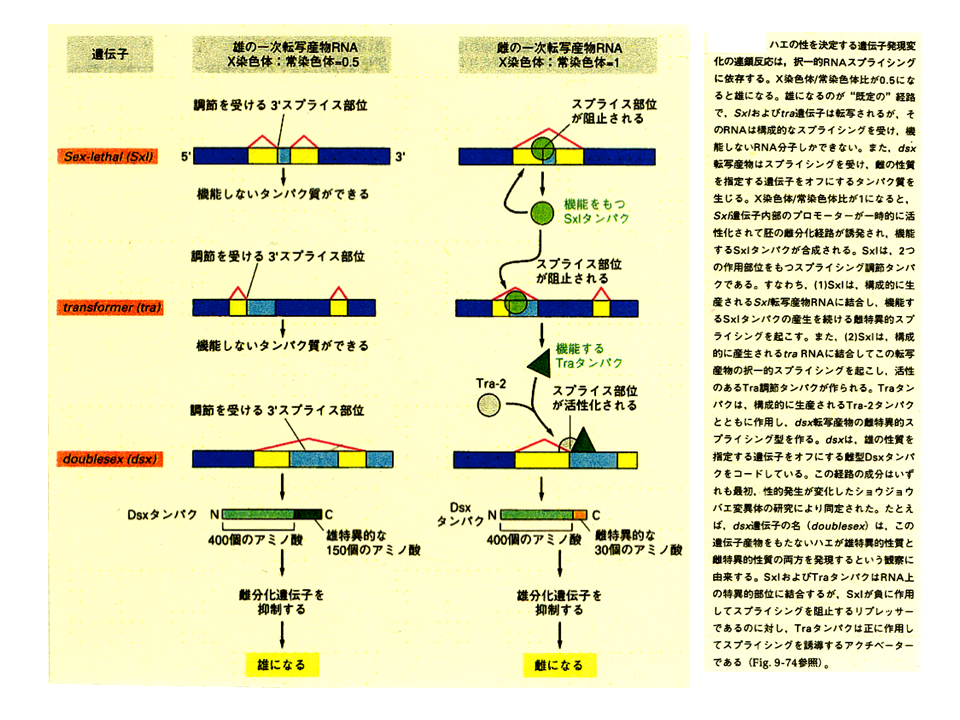
選択的スプライシングは非常に大きな働きをしており、ショウジョウバエでは性の決定に関係している。染色体がXXであれば雌、XYであれば雄といとも簡単に性が決まっているようにみえるが、いろいろな遺伝子が複雑に働いて性が決まるのである。例えば、Sex-lethal、transformer、doublesex遺伝子は、選択的なスプライシングによって、機能する蛋白質を生み出したり、機能しない蛋白質を生み出したり、機能の異なる蛋白質を生み出したりしている。

説明文の4
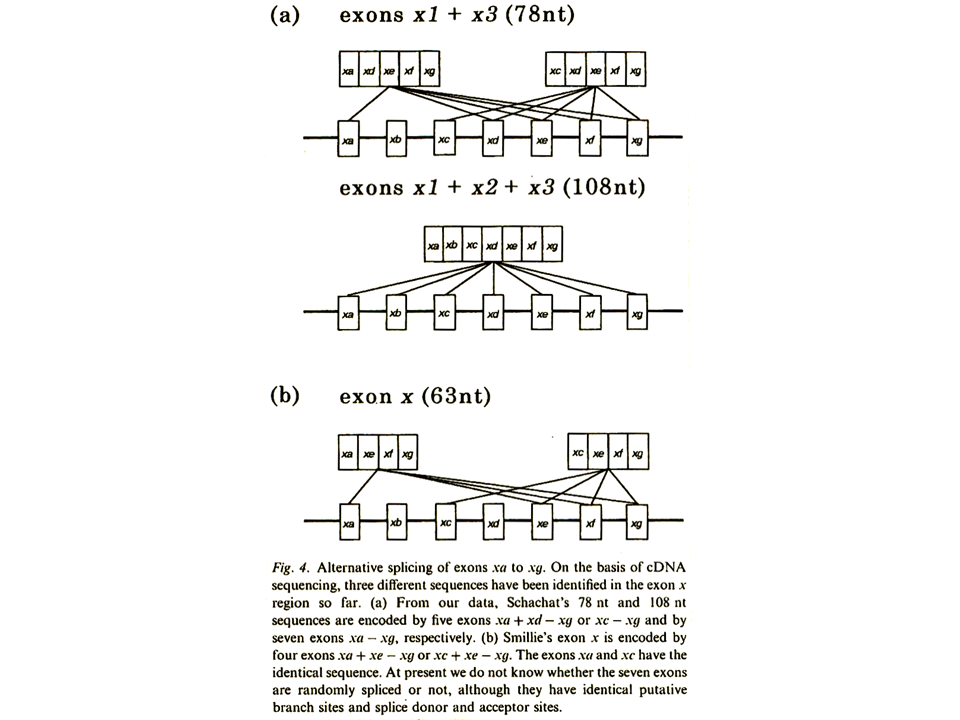
当初速筋型トロポニンT遺伝子のエクソンx領域は, mRNAの塩基配列から、x1、x2、x3の3つのエクソンから成るとされていた。しかし、遺伝子の構造を決定したところ、xaからxgの7つのエクソンが含まれることが明らかになった。エクソンの構造は遺伝子自身の構造を決定しないと明らかにならない。
説明文の5
当初mRNAの塩基配列から見出されたエクソンx1+x2+x3は実際にはxa+xb+xc+xd+xe+xf+xgであり、エクソンx1+x3は、xa+xd+xe+xf+xgまたはxc+xd+xe+xf+xgであることがわかった。xaとxcは全く同じ配列なので区別できない。また、当初エクソンxとされていたものはxa+xe+xf+xgまたはxc+xe+xf+xgであることがわかった。
説明文の6
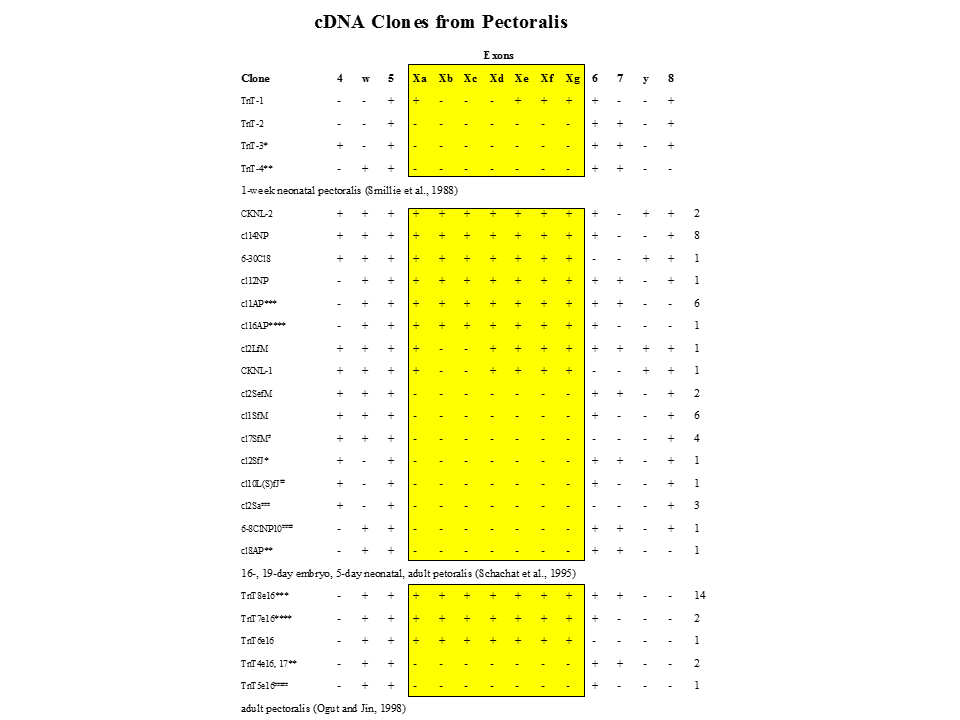
一方、実際にどのような選択的スプライシングが行われているかは、mRNAの塩基配列を決定しないと明らかにならない。ニワトリの大胸筋では、128とおりのパターンのうちxa+xb+xc+xd+xe+xf+xg、xa+xd+xe+xf+xg(あるいはxc+xd+xe+xf+xg)、xa+xe+xf+xg(あるいはxc+xe+xf+xg)、すべてのエクソンを含まない、の4(6)とおりのパターンしかみつからなかった。
説明文の7
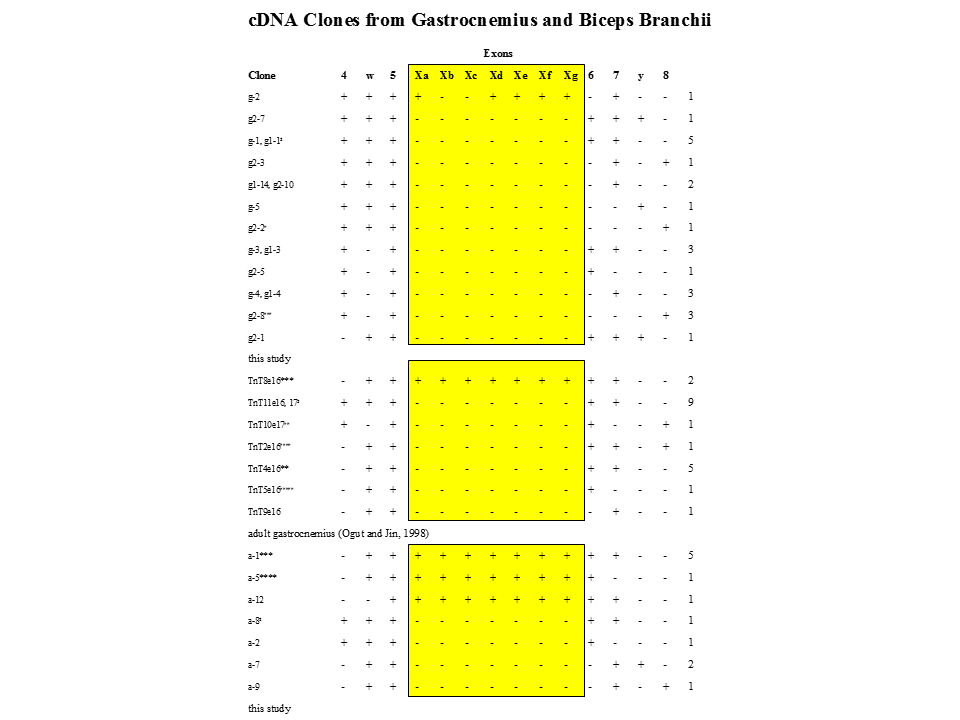
上腕と頭筋においても、xa+xb+xc+xd+xe+xf+xg、xa+xd+xe+xf+xg(あるいはxc+xd+xe+xf+xg)、すべてのエクソンを含まない、の3(5)とおりのパターンしかみつからなかった。それゆえ、すべての筋肉を発生に伴って調べてはいないが、選択的スプライシングは制御されていると思われる。
説明文の8
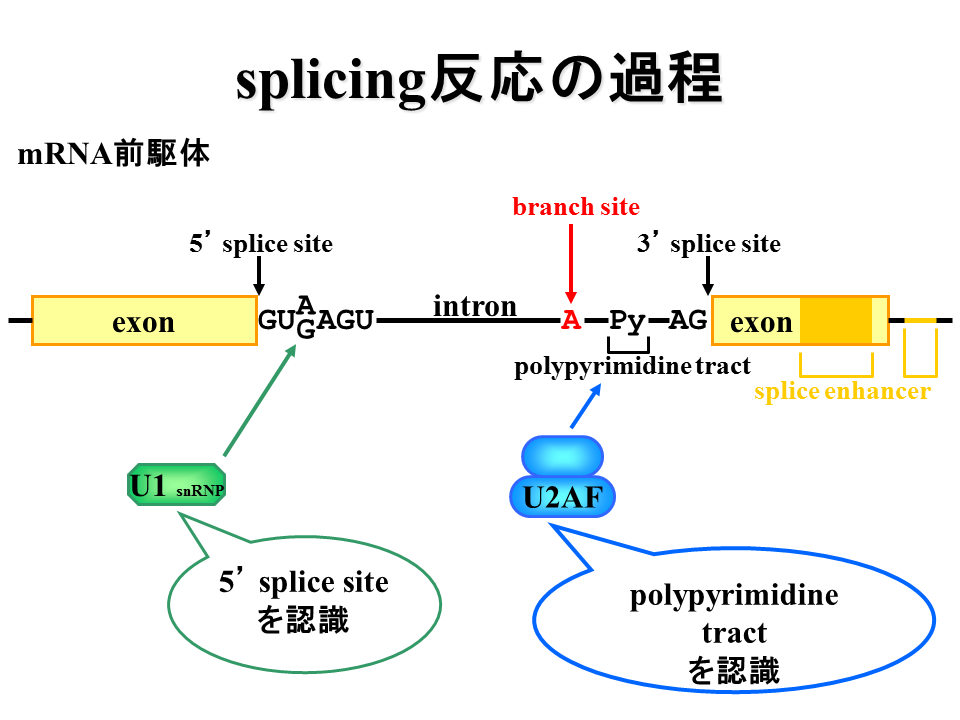
エクソンx領域で選択的スプライシングはどのように制御されているのだろうか?スプライシング制御には5‘splice site、3’spliced site、polypyrimidine tract、branch site、エクソンの中あるいは周辺に存在するsplice enhancerが重要だとされている。
説明文の9
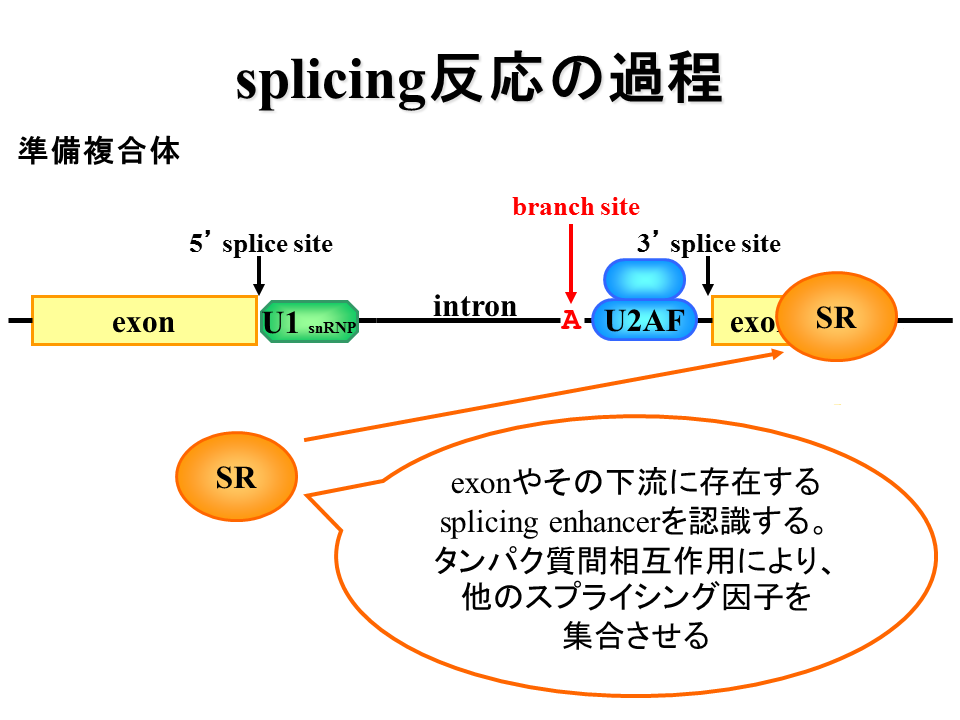
U1蛋白質が5‘splice siteに結合し、U2AF蛋白質がpolypyrimidine tractに結合する。また、SR蛋白質がエクソン周辺に結合する。
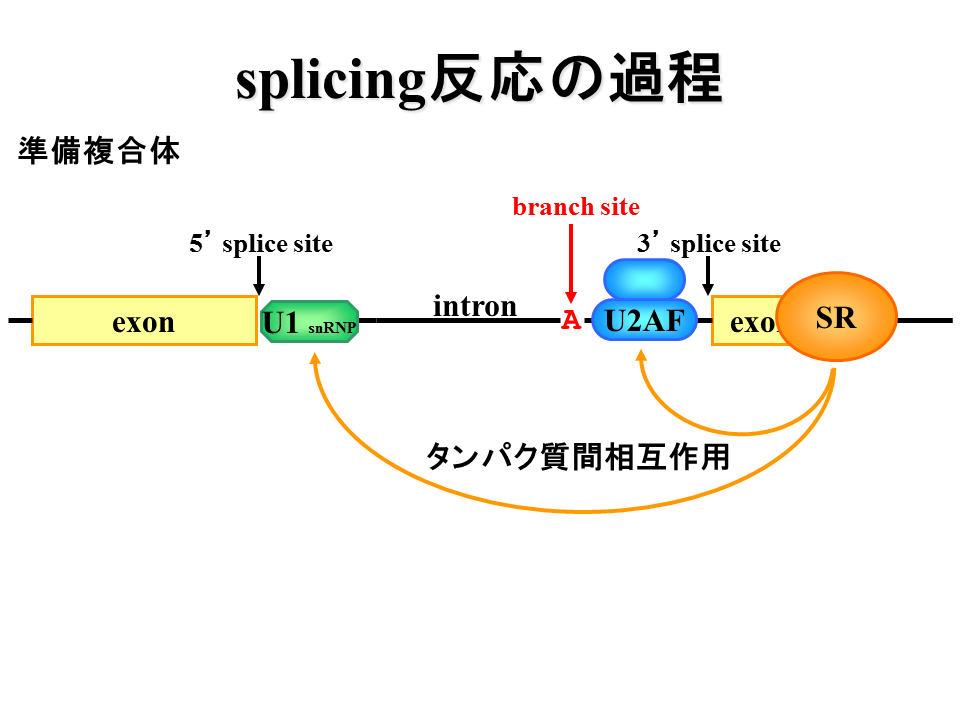
説明文の10
SR蛋白質がU1蛋白質とU2AF蛋白質と相互作用する。
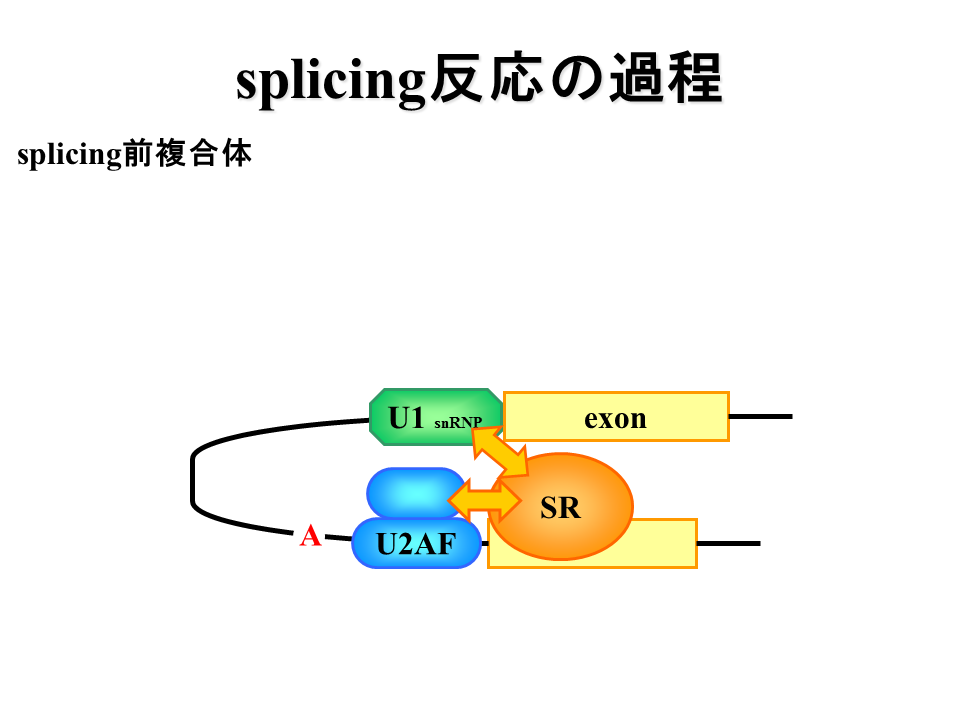
説明文の11
エクソン同士が近づくように折り畳まれる。
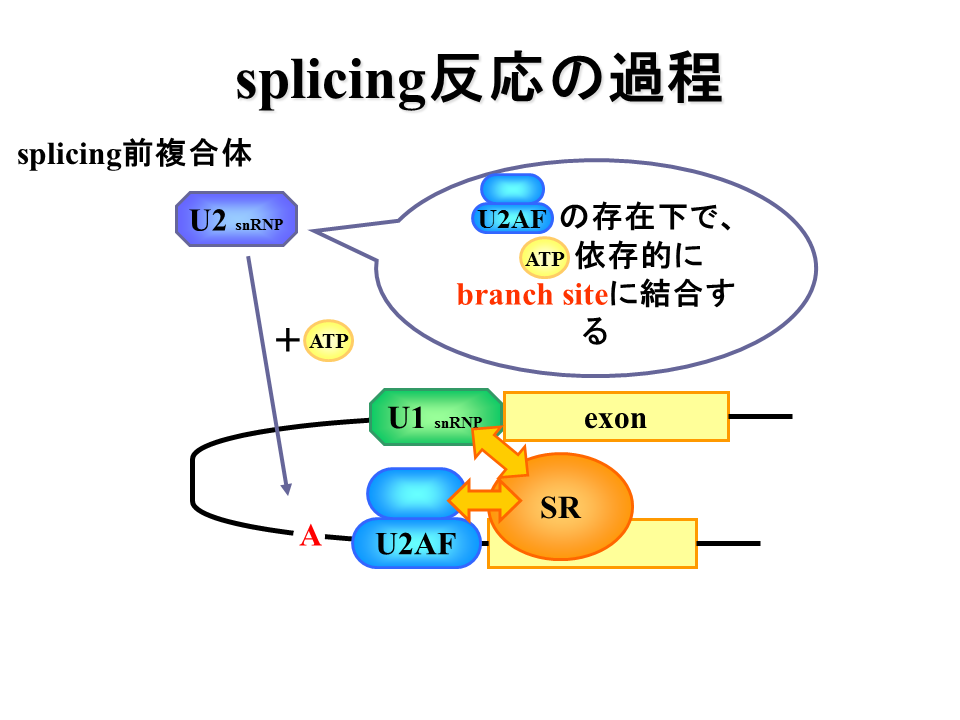
説明文の12
U2蛋白質がbranch siteに結合する。
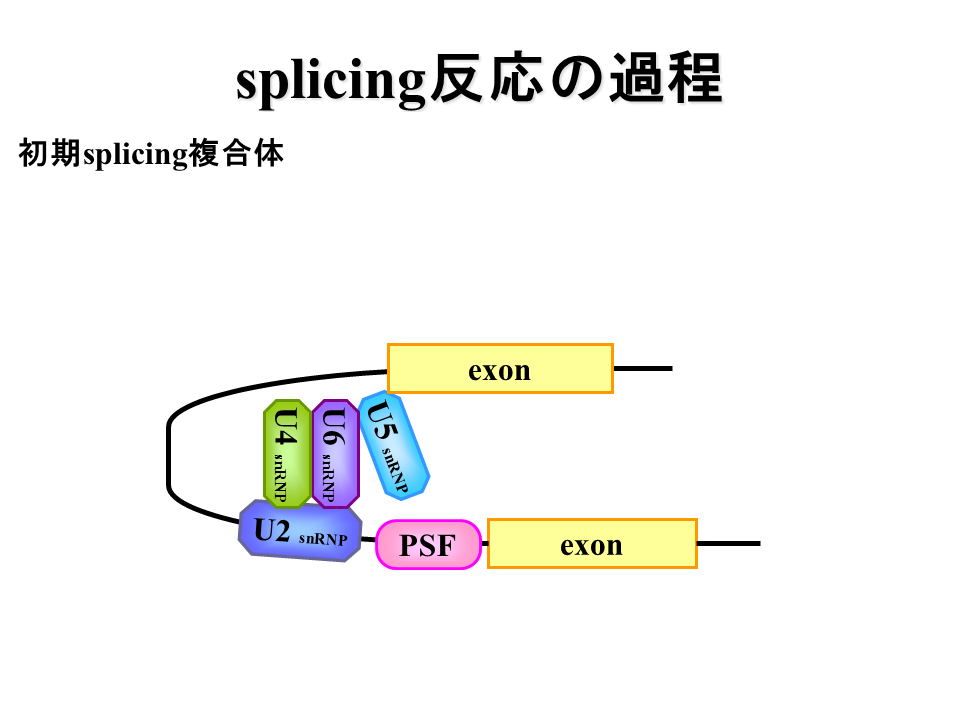
説明文の13
U2AF蛋白質がPSF蛋白質に置き換わり、U4、U5、U6蛋白質が加わる。
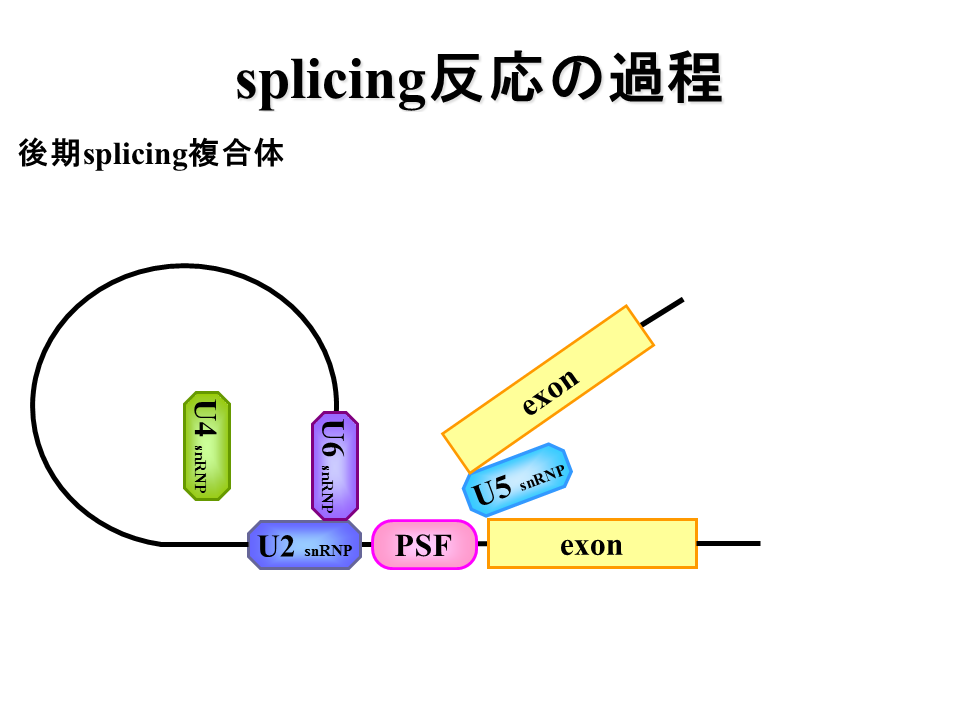
説明文の14
U2とU6蛋白質の相互作用によって、投げ縄状構造ができる。
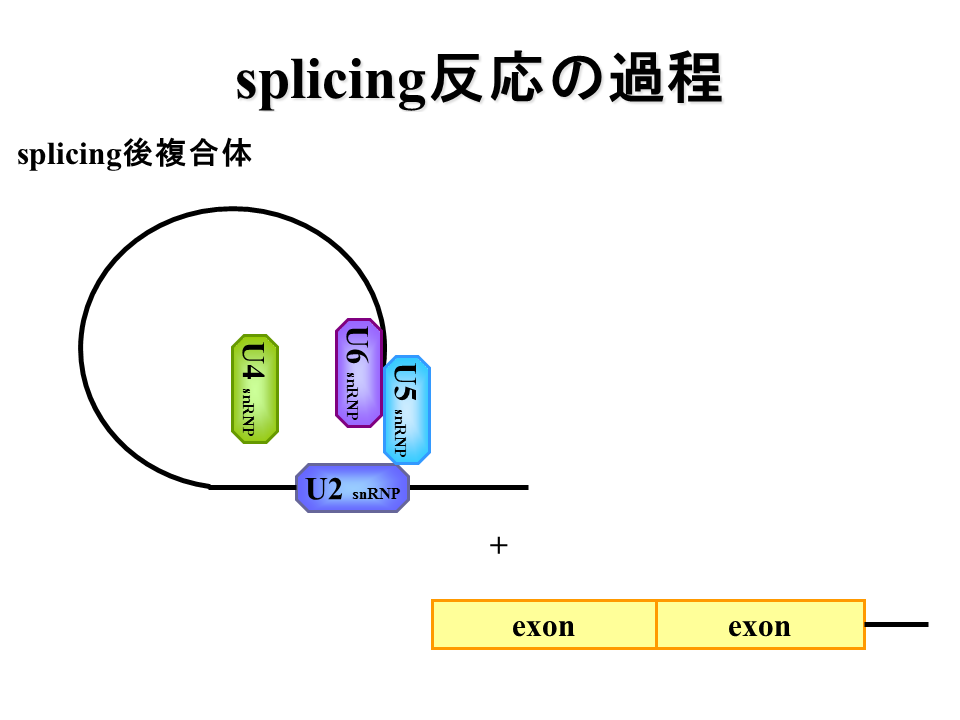
説明文の15
イントロン部分が切り出され、エクソン同士がつながる。

説明文の16
それではエクソンx領域で、スプライシングの制御に重要な部分はどうなっているだろうか。3’splice site(Splice acceptor)はすべてのエクソンでagであり、5’splice site(Splice donor)はすべてのエクソンでgtであり、違いはなかった。Branch siteもすべてのエクソンでtcctaacで違いがなかった。polypyrimidine tractは多少塩基配列の違いがあり、長さも微妙に異なるが、大きな違いはなかった。エクソン自体の配列もxgが3塩基長いだけで、大きな違いはなかった。ほとんど違いがないのに、なぜスプライシングのパターンが128とおりでなく、4あるいは6とおりしかないのであろうか?今後の研究課題である。
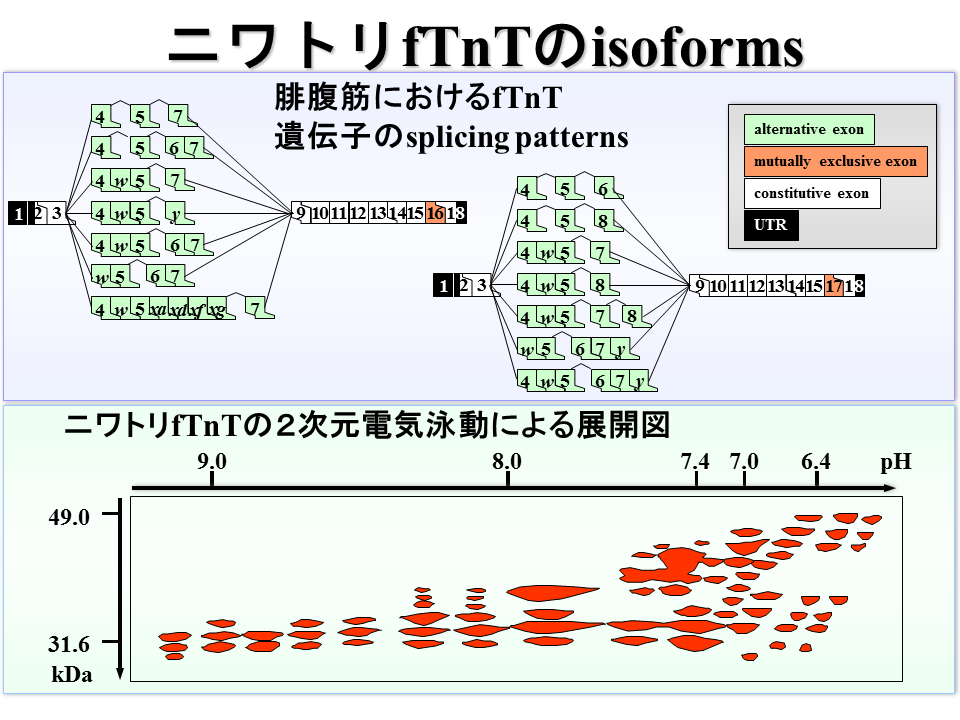
説明文の17
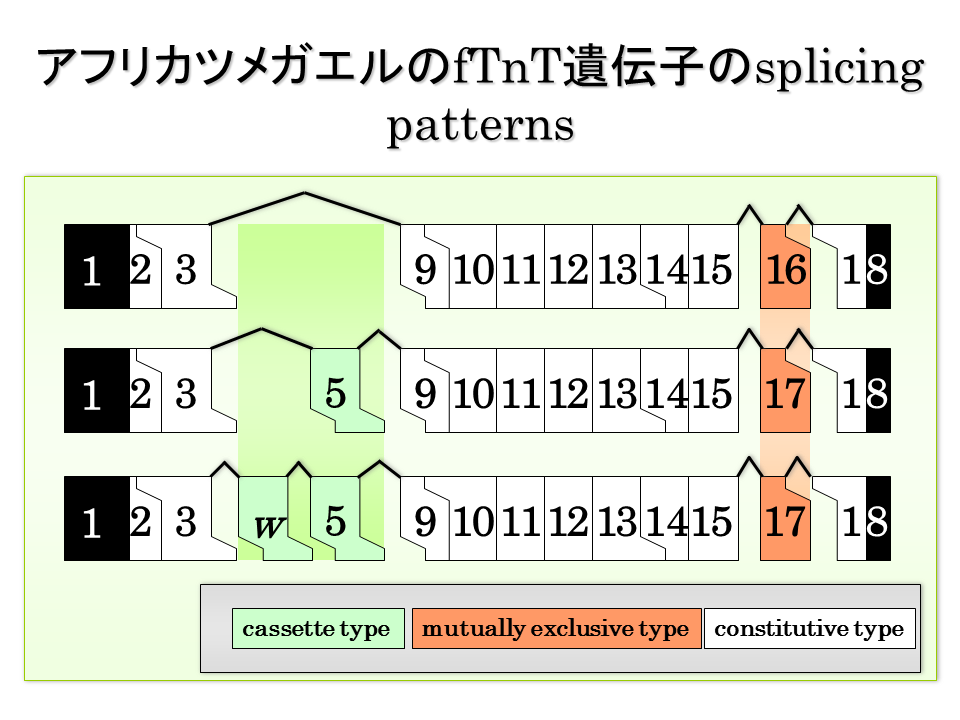
今度は速筋型トロポニンT遺伝子の相互排他的エクソンについて考えてみる。エクソンは16と17は、どちらかが必ず選択される。腓腹筋のmRNAの塩基配列を決定すると、エクソン4から8までは様々なパターンで選択されるが、16と17はどちらか一方が選択される。

説明文の18
混合筋の腓腹筋では、基部(proximal)でエクソン16と17はどちらも選択されたが、先端部(distal)ではエクソン17だけが選択された。一方、速筋の大胸筋ではエクソン16と17どちらも選択され、遅筋の前広背筋ではどちらも選択されない(遅筋なので速筋型トロポニンTは発現しない)。
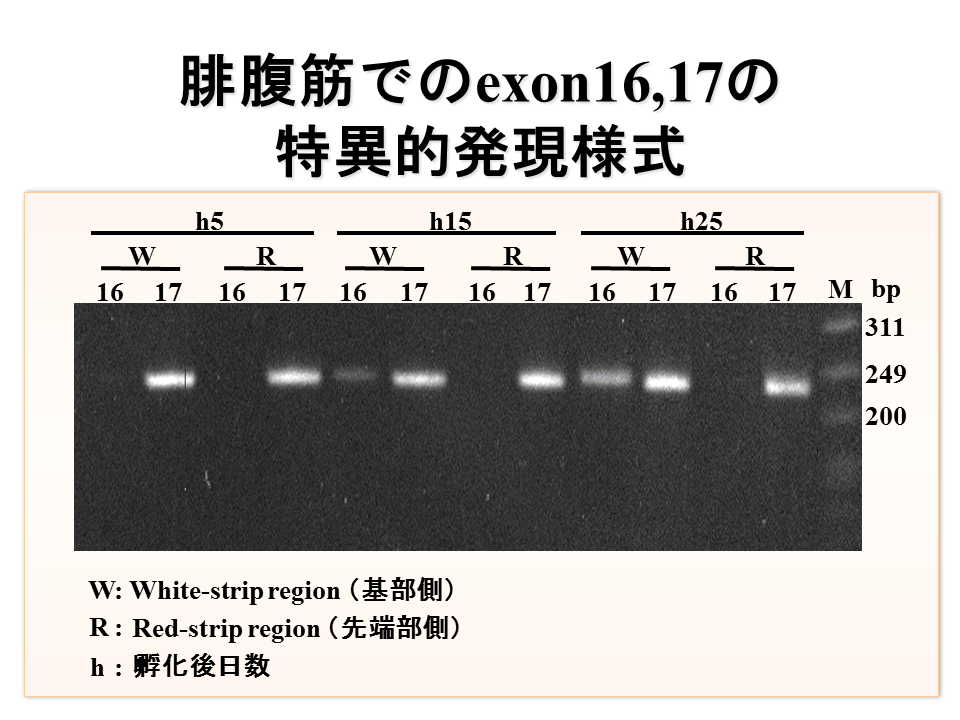
説明文の19
腓腹筋ではエクソン16と17の選択は発生段階により変化した。孵化後5日目では、基部でも先端部でもエクソン17のみが選択されたが、基部では孵化後15日目、25日目と発生段階が進むに連れエクソン16が選択されるようになった。一方、先端部では変化がなく、発生を通してずっとエクソン17のみが選択された。
説明文の20
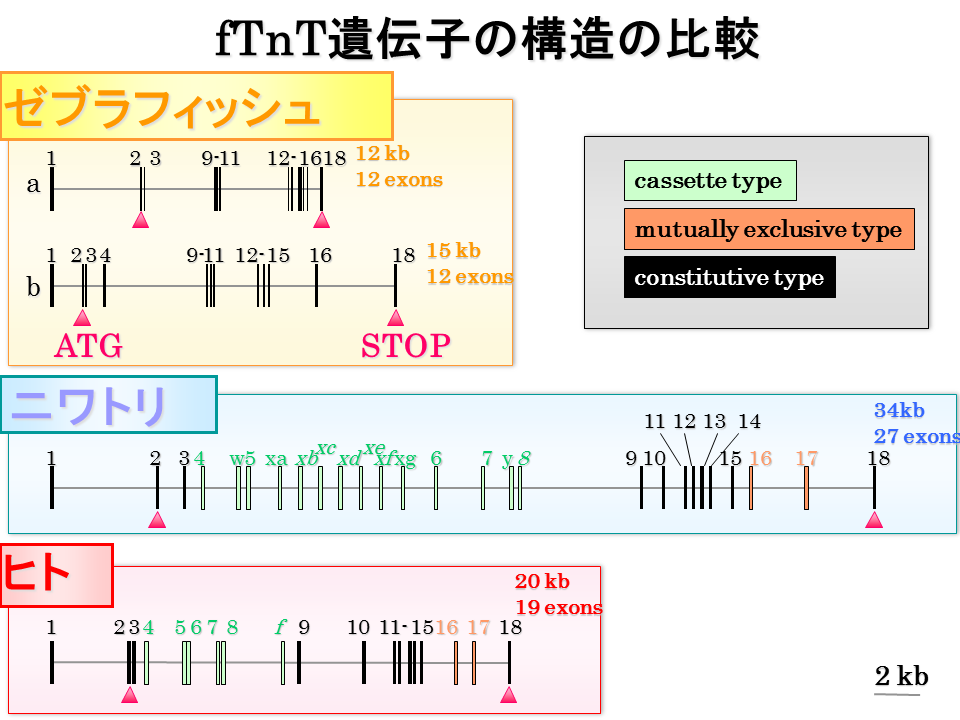
ニワトリとヒトでは、エクソン16と17が相互排他的に選択される。しかし、ゼブラフィッシュでは、選択的なエクソンがない。そこで、選択的されるエクソンと、されないエクソンの違いを探ってみた。
説明文の21
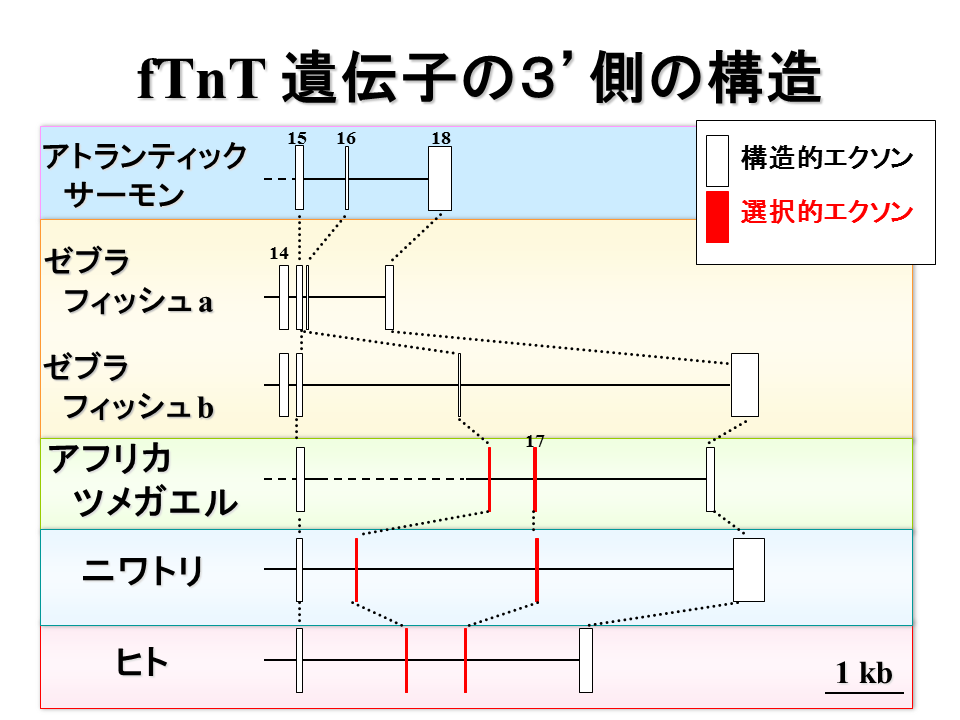
アトランティックサーモンでも選択されるエクソンがなかったので、硬骨魚類では選択的スプライシングが、この領域では進化していないのかもしれない。一方、アフリカツメガエルでは、エクソン16と17ガ存在した。
説明文の22
塩基配列を決定したmRNAは少ないが、エクソン16と17な相互排他的に含まれていた。したがって、選択的なエクソン16と17は陸上動物で進化したのかもしれない。

説明文の23
エクソン16と17の関係を調べると、アトランティックサーモンやゼブラフィッシュのエクソン16(16番目のエクソン)は陸上動物のエクソン16に類似した。
説明文の24
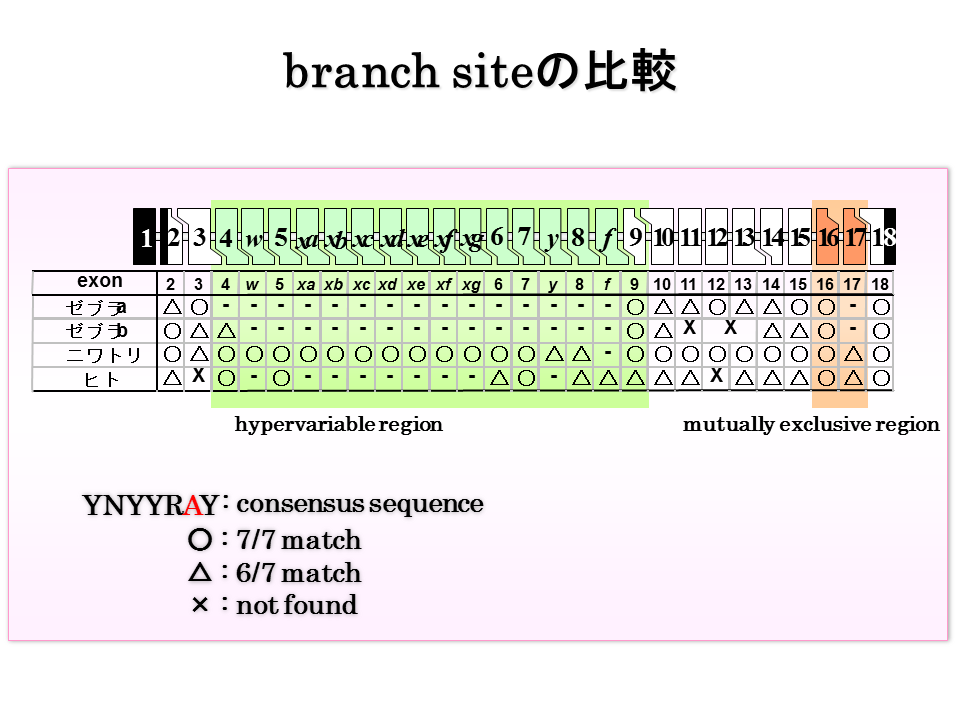
エクソン16と17のブランチサイト(branch site)を比べてみた。tcctaacかそれとは1塩基だけ異なるブランチサイトは概ねどのエクソンにもあるようだ。-はその動物に相当するエクソンがないことを示す。

説明文の25
ブランチサイトの数を比べてみた。配列がtcctaacでの場合白丸で、それとは1塩基だけ異なる時は黒丸で示した。硬骨魚類では、ゼブラフィッシュaは情報が不足しているが、ブランチサイトの数は多いようにみえる。陸上動物のエクソン16と17の間で、ブランチサイトの数にはいくらかの違いがある(エクソン16の方が多い)ようだが、生物種による違いなのかもしれない。
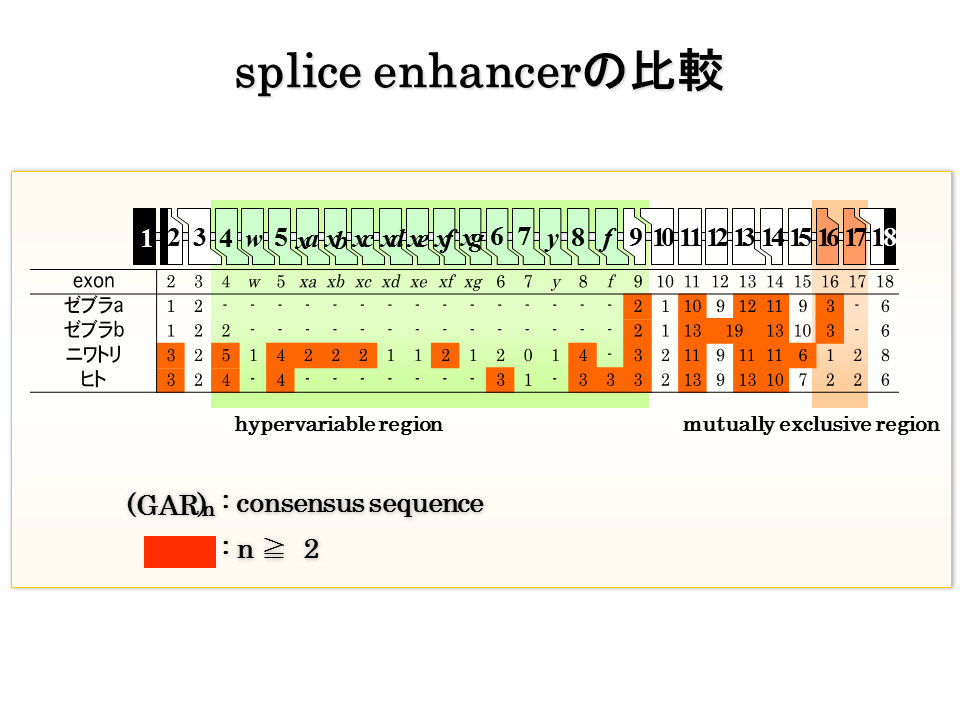
説明文の26
スプライシングを活性化するスプライスエンハンサー(splice enhancer)数は、硬骨魚類でいくらか多いようであるが、陸上動物のエクソン16と17を含めて明確な違いがあるようにはみえない。
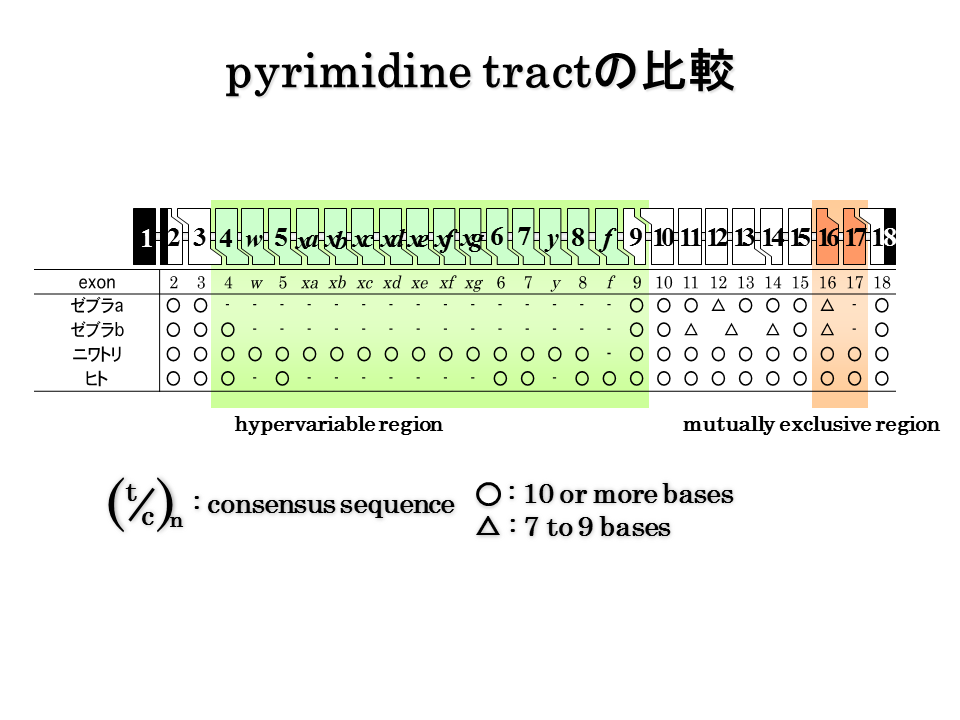
説明文の27
ピリミジントラクト(polypyrimidine tract)も、硬骨魚類でいくらか短いようであるが、陸上動物のエクソン16と17を含めて明確な違いがあるようにはみえない。
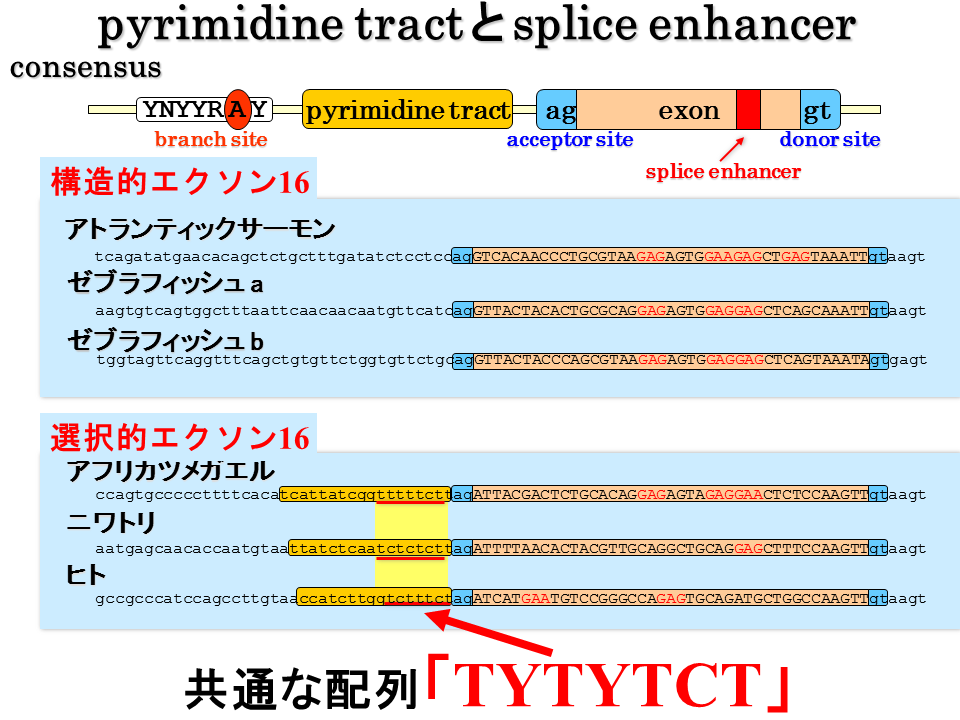
説明文の28
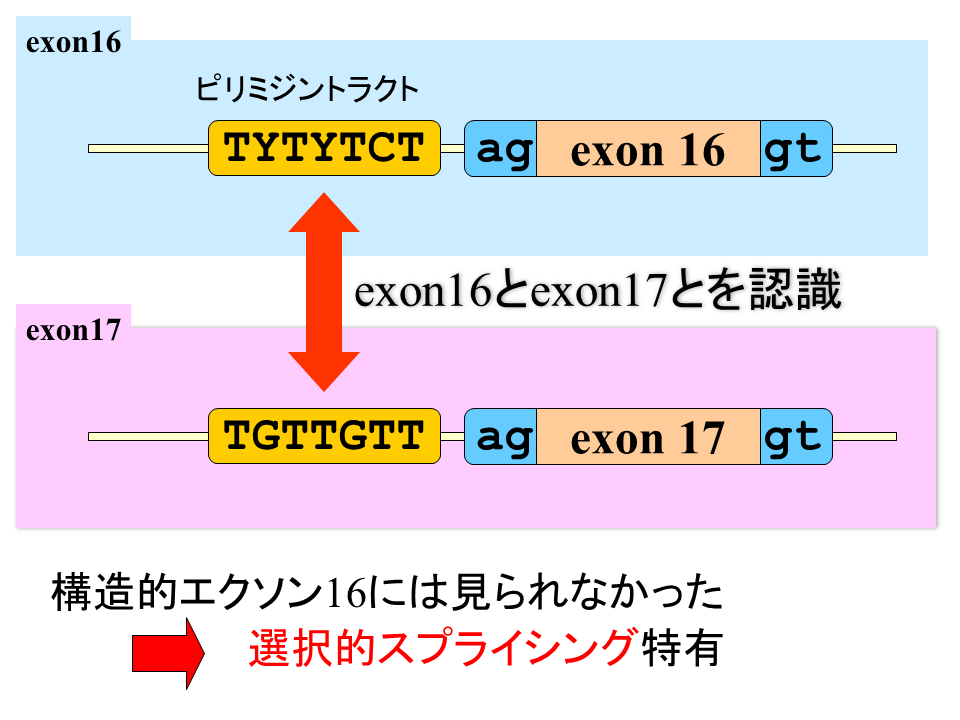
ピリミジントラクトの塩基配列を詳細に調べてみると陸上動物の速筋型遺伝子のエクソン16の間には、tytytct(yはtかcを示す)で表される共通の配列がある。選択的スプライシングが行われない硬骨魚にはこれに一致する配列はない。
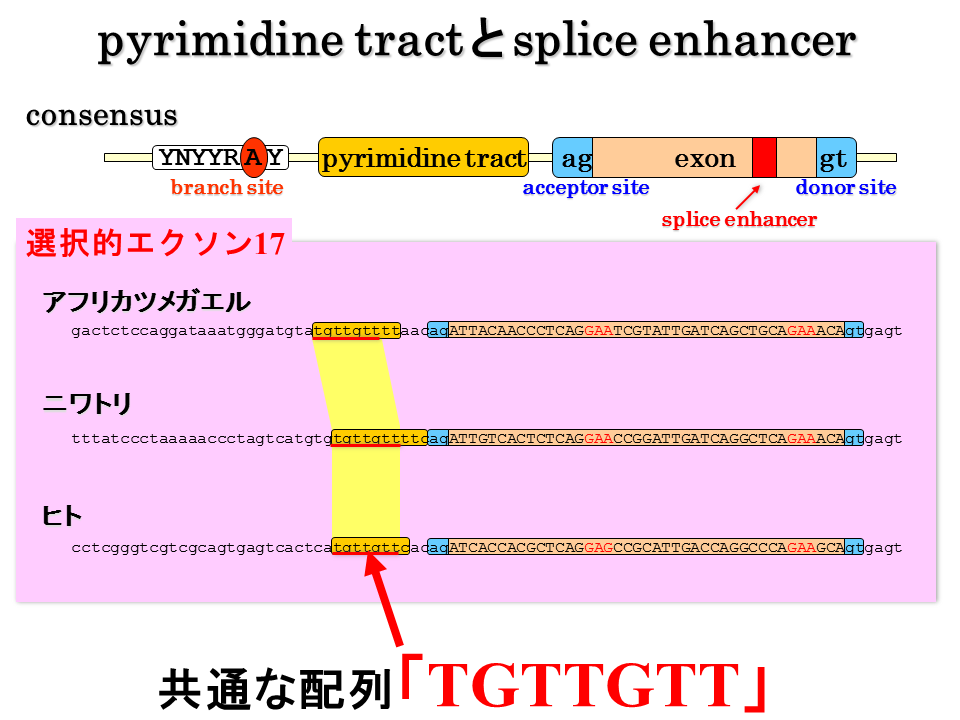
説明文の29
一方、硬骨魚にはない選択されるエクソン17には、ピリミジントラクトにtgttgttという共通の配列がある。エクソン16と17にみられるこれらの配列が、選択的スプライシングをするか否か、エクソン16と17のどちらを選択するか、を決定するシグナルではないか?

説明文の30
以上をまとめると、硬骨魚類では相互排他的な選択的スプライシングは行われず、おそらく両性類以上の陸上動物でエクソン16と17の相互排他的な選択的スプライシングが行われる。したがって、魚類のエクソン16にはなく、陸上動物のエクソン16と17にみられるものが、選択的スプライシングの機構を解く鍵だと思われる。、魚類のエクソン16と陸上動物のエクソン16と17の間では、明確な差がみられなかったが、陸上動物のエクソン16と17のピリミジントラクトには各々に特異的な配列が存在することがわかった。
説明文の31
ピリミジントラクトに存在す特異的な配列によって、選択的なスプライシングが起こるのであろうか?また、エクソン16と17の選択における区別が起こるのであろうか?今後の課題である。
説明文の32
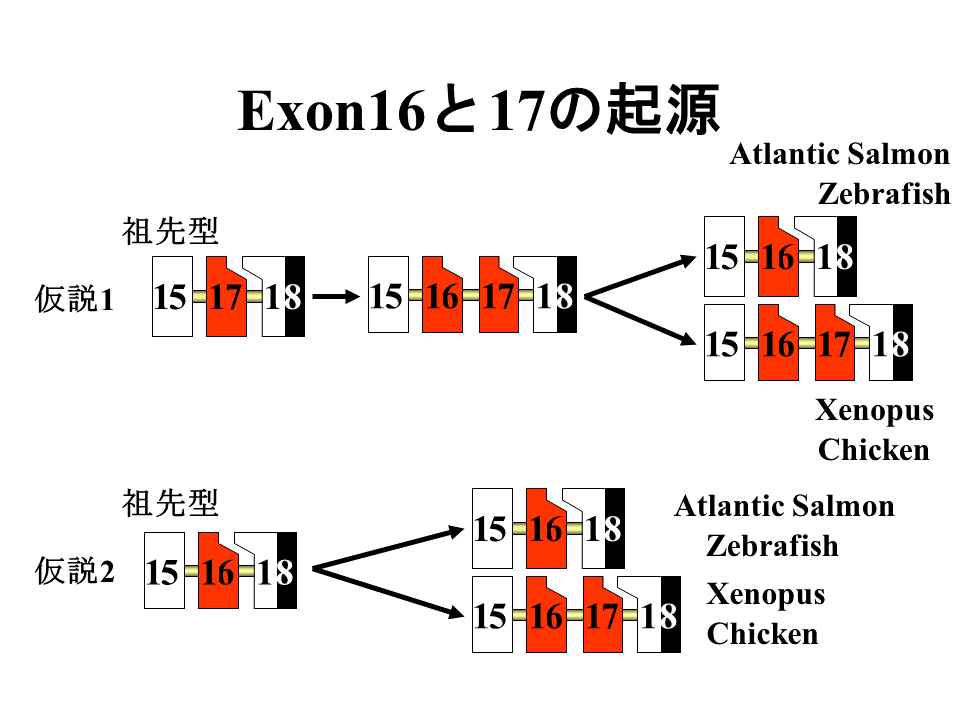
エクソン16と17において、祖先型はどちらなのであろうか?祖先型は17で、それがエクソンの重複で16と17になり、硬骨魚では16だけが残り、陸上動物では両方が残ったという仮説1、祖先型は16で、硬骨魚ではそのまま16だけをもつが、陸上動物ではエクソンの重複によって16と17になったという仮説2、を考えた。

説明文の33
速筋型、遅筋型、心筋型トロポニンT遺伝子は1つの遺伝子から進化したといわれる。遅筋型と心筋型トロポニンT遺伝子のエクソン16にあたるエクソンは、速筋型のエクソン17に類似する。それゆえ、エクソン17が祖先型ではないかと思われる。このことは仮説1を支持している。
説明文の34
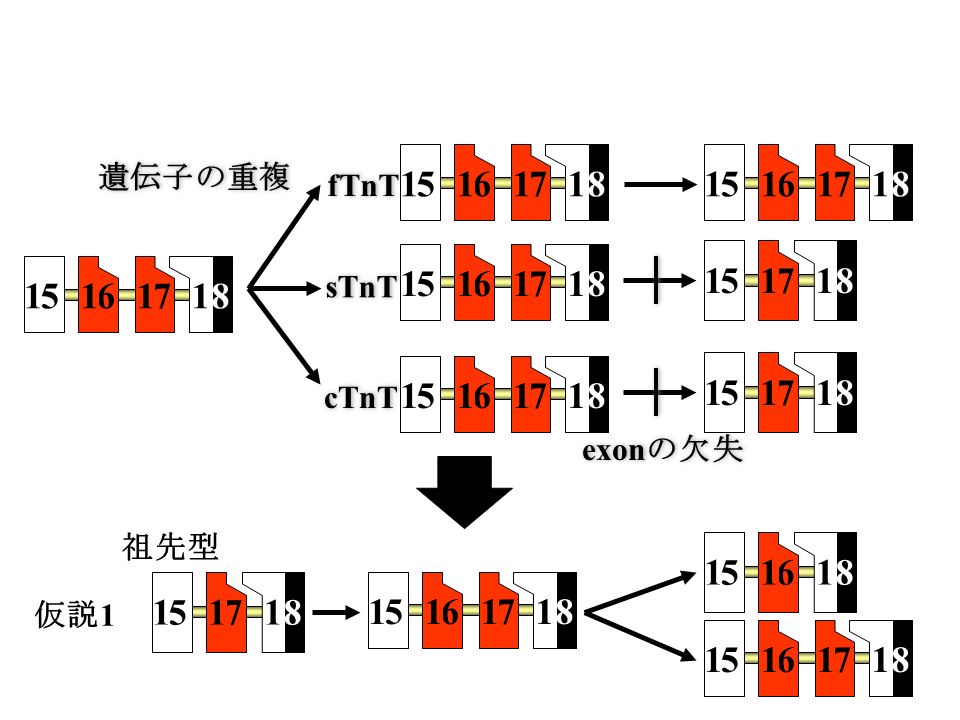
しかし、速筋型、遅筋型、心筋型トロポニンT遺伝子に分かれる前の祖先型遺伝子が、既にエクソン16と17をもっており、遅筋型と心筋型遺伝子でエクソン16が失われたと考えると、エクソン16と17の祖先は16でも17でもどちらでもよくなってしまう。
説明文の35
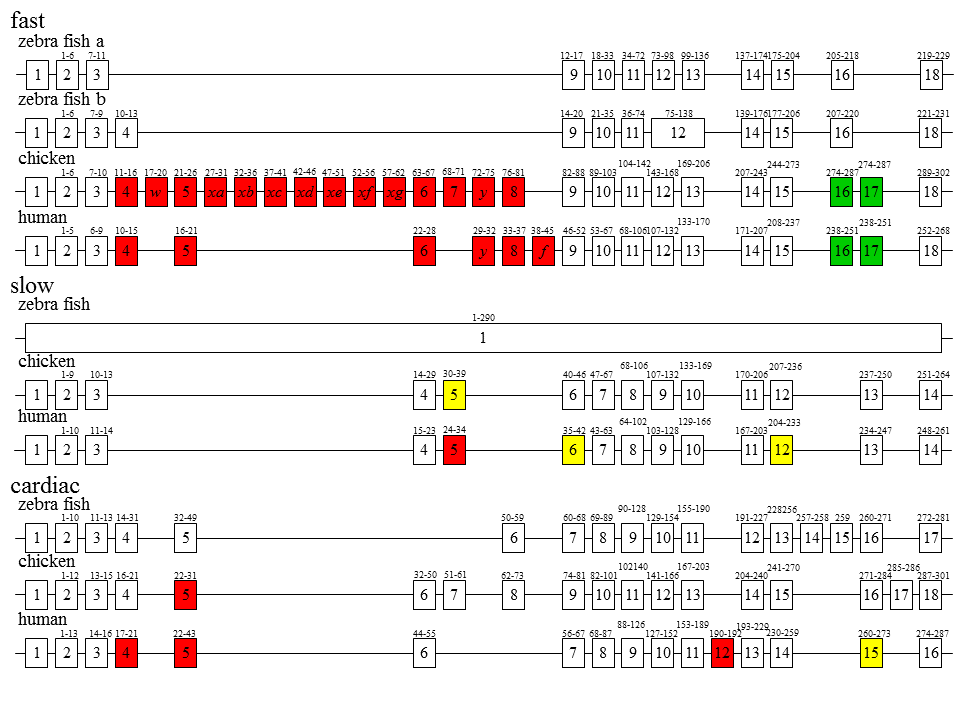
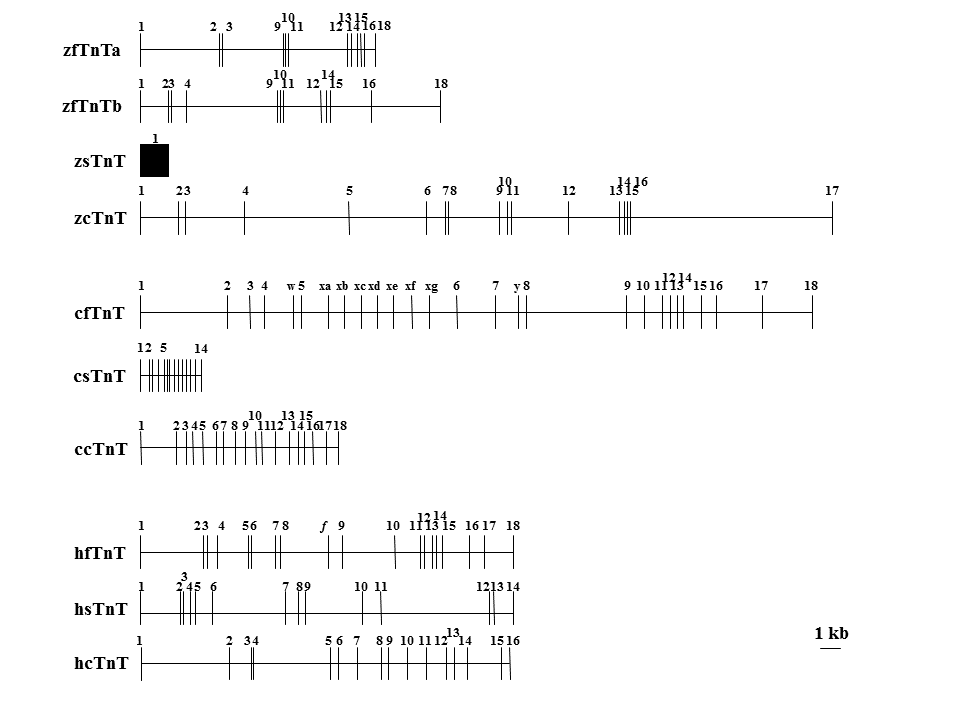
そもそも速筋型、遅筋型、心筋型トロポニンT遺伝子は本当に1つの遺伝子から進化したのだろうか。ゼブラフィッシュ(zc)やにわとり(c)では、3つの遺伝子の構造が大きく異なるようにみえる。一方ヒト(h)では、3つの遺伝子の構造(遺伝子の長さやエクソンの数)は比較的似ているようにみえる。
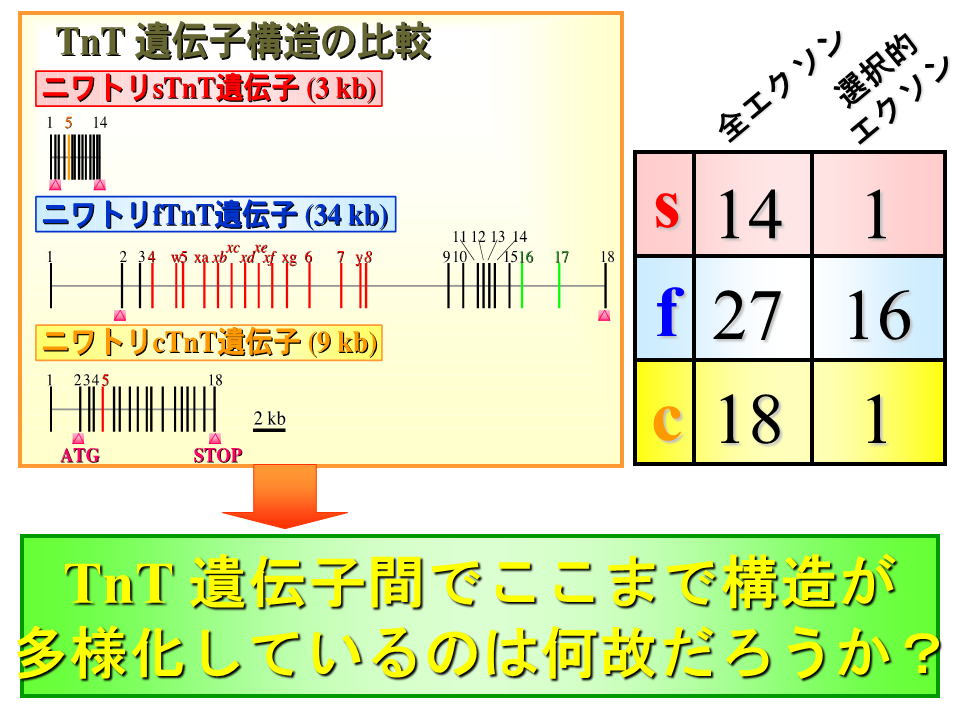
説明文の36
ニワトリの遺伝子では、3つの遺伝子の間で遺伝子の長さやエクソンの数が大きく異なる。
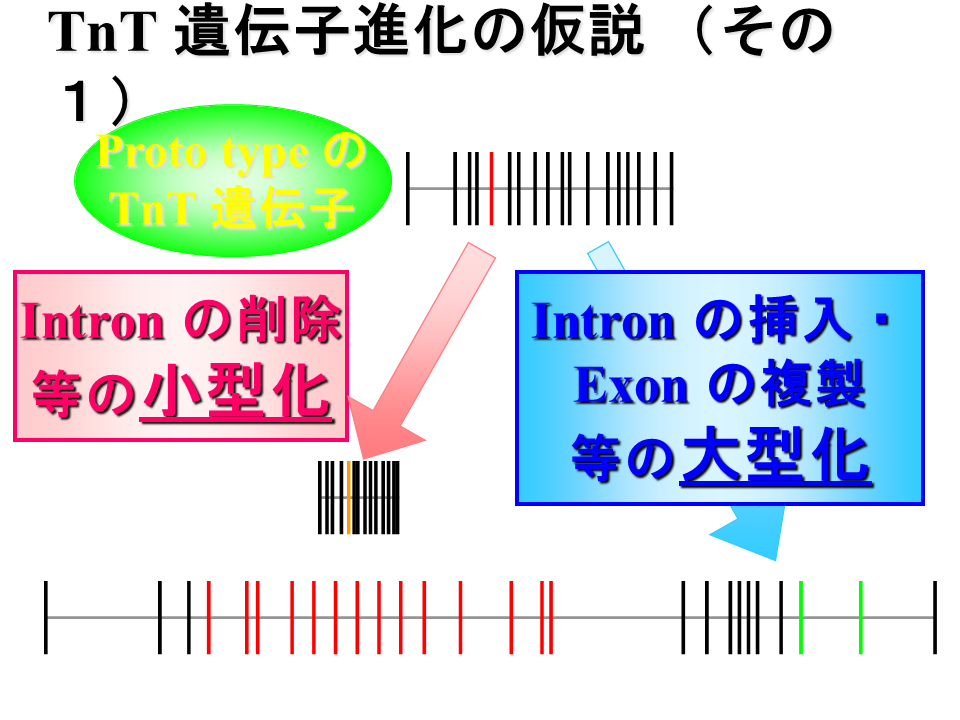
説明文の37
遺伝子の進化を考えてみる。ヒトの遺伝子のように約10 kbくらいの心筋型遺伝子のようなものが祖先だとすると、心筋型遺伝子はほぼそのまま(エクソンの数も速筋型と遅筋型の中間)であり、遅筋型遺伝子では極端な小型化が、速筋型遺伝子では極端な大型化が起こったと考えられる。
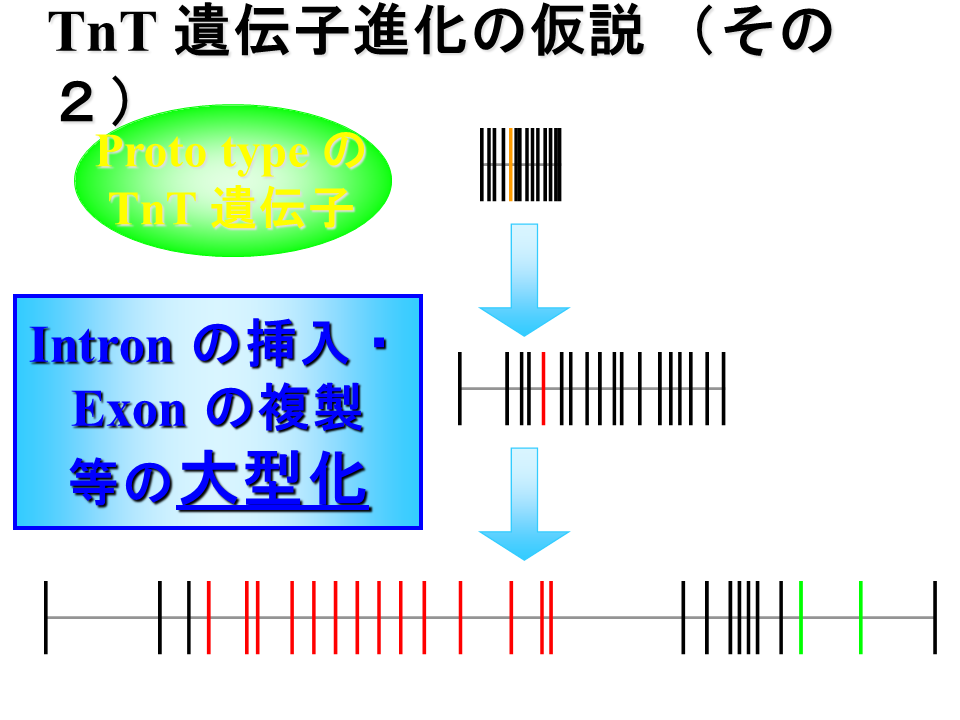
説明文の38
一方、一番短い遅筋型の遺伝子のようなものが祖先型で、遅筋型はそのまま、大型化して心筋型、さらに速筋型になったとも考えられる。もちろん、この遺伝子進化の大型化の逆で、速筋型遺伝子のような大きな遺伝子を祖先として遺伝子進化の小型化が起こった可能性もある。