説明文の1
遅筋型トロポニンT遺伝子の発現調節には、E-box4が重要らしいことがわかってきた。それでは、速筋型遺伝子、心筋型遺伝子ではどうだろうか?それについては既に報告がある。

説明文の2
速筋型トロポニンT遺伝子、遅筋型トロポニンT遺伝子、心筋型トロポニンT遺伝子の上流域の塩基配列である。赤字で示したものは遺伝子発現の調節にかかわりそうな配列である。遅筋型遺伝子だけでなく、他の遺伝子の上流にも複数のcis-acting elementの候補がある。

説明文の3
トロポニンT遺伝子の中で、一番最初に分析されたのは、心筋型遺伝子であり、ニワトリの遺伝子ではM-CAT(CATTCCT)が重要であることが既に記載されていた。速筋型ではどうであろうか?
説明文の4
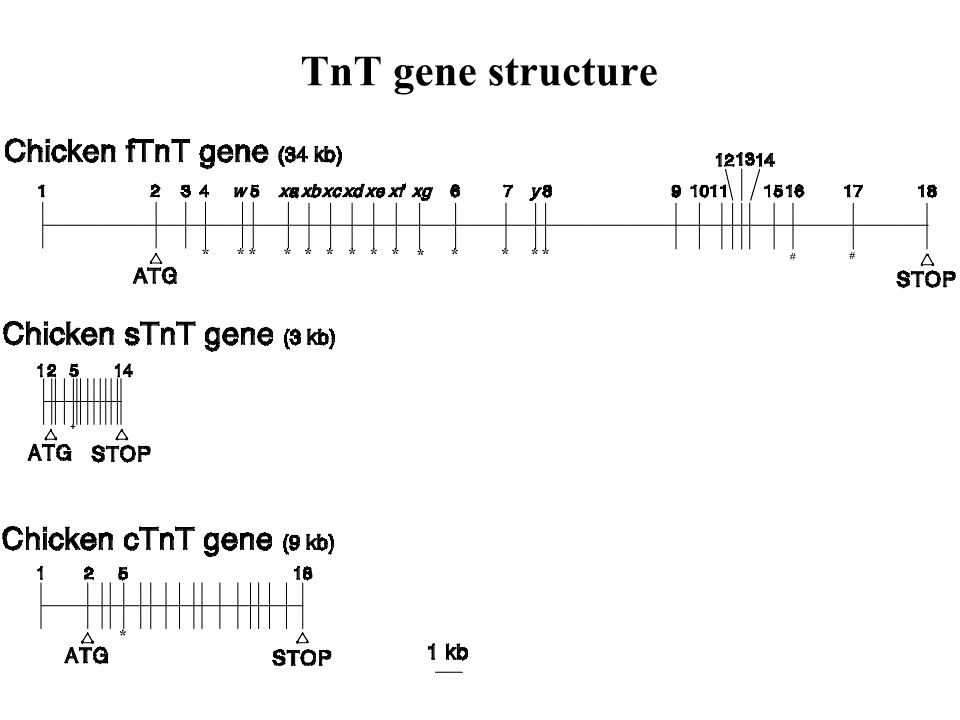
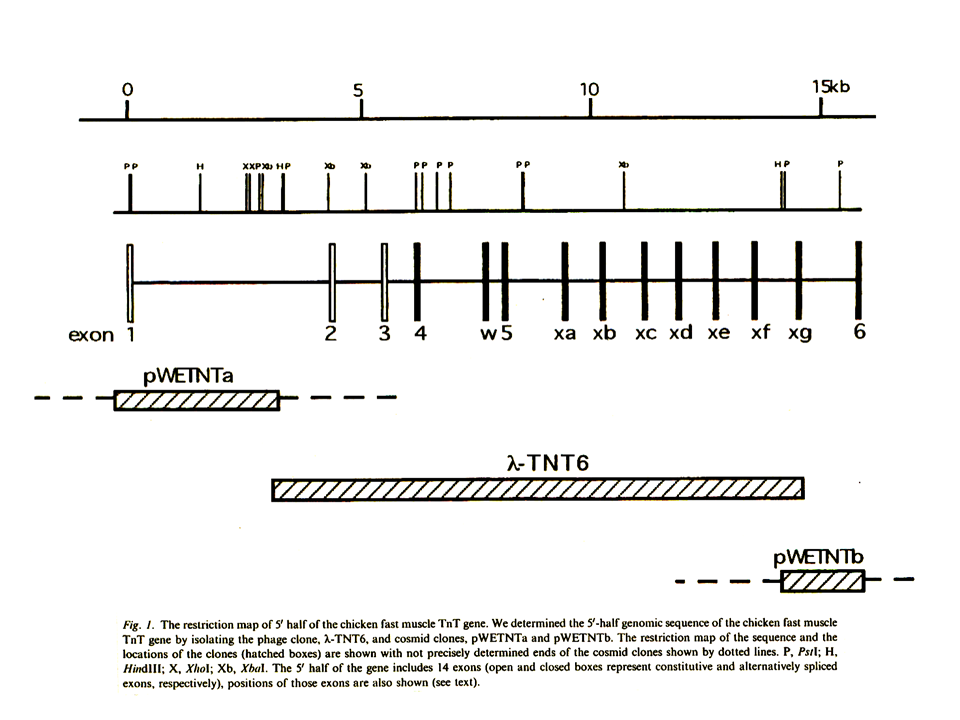
速筋型遺伝子については、まず前半部分(エクソン6まで)の構造を決定した。

説明文の5
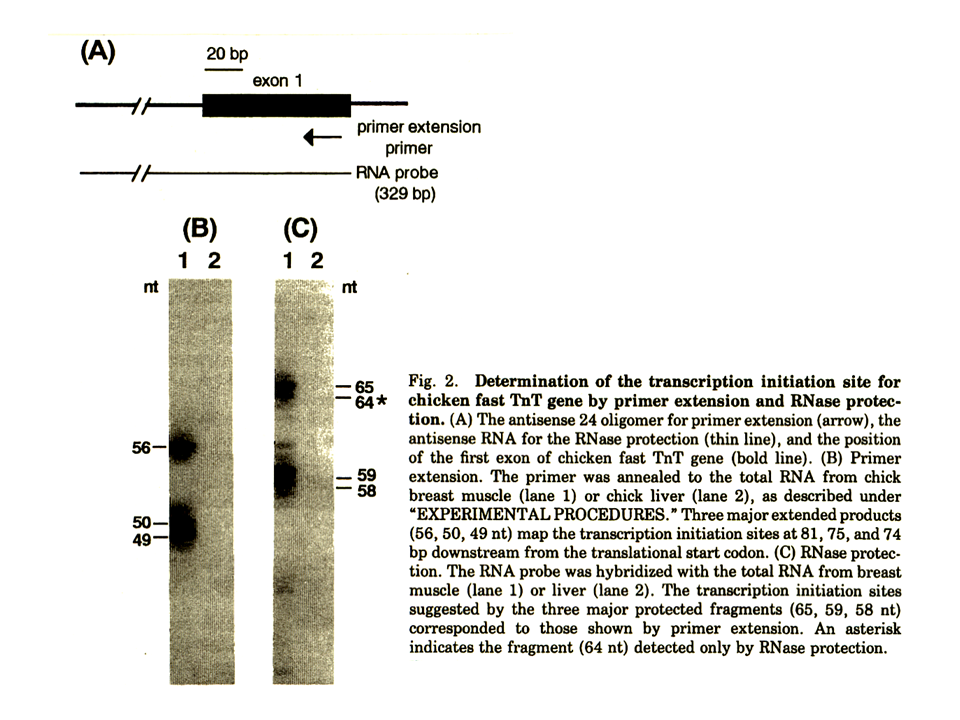
その塩基配列を示す。赤字の部分がエクソンである。遺伝子の端(mRNAの始まり)あるいはエクソン1の先端は、遅筋型遺伝子と同様にprimer extensionによって決定した。
説明文の6
primer extensionでは、56、50、49 ntにバンドがみられたが、mRNAの先端が何らかの原因で不完全である可能性もあるため、一番サイズの大きいバンドが遺伝子の端(mRNAの始まり)を示していると考えた。また、RNA protectionによっても遺伝子の端(mRNAの始まり)を調べた。遺伝子の塩基配列から端(mRNAの始まり)のありそうな部分のRNAプローブを作製した。これをmRNAと結合させると2本鎖になるが、1本鎖だけを切断する酵素でプローブとmRNAが結合していない部分を切断した。2本鎖となっており酵素によって切断されなかった最もサイズの大きいバンドは65と64 ntであった。64 ntのバンドが示す位置はprimer extensionの示す位置と同じであったため、ここを遺伝子の端(mRNAの始まり)、すなわち+1、とした。1では、速筋の大胸筋から得られたmRNAを用いたが、2では肝臓から得られたmRNAを用いた。肝臓では、トロポニンTが発現されていないので、バンドは全くみられなかった。

説明文の7
上流域には、E-box 1と2やM−CAT(CATTCCT)と1塩基異なるM-CAT-like(CACTCCT)等がみられた。
説明文の8
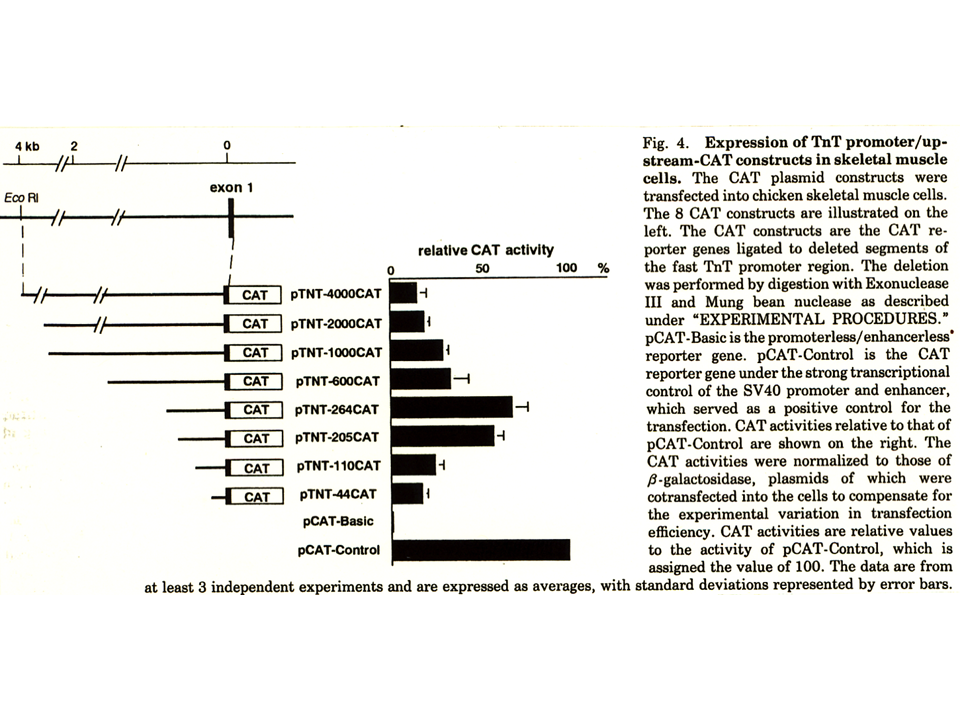
速筋型遺伝子の上流を徐々に削って、CAT遺伝子の上流に連結し、CAT assayを行った。その結果、上流264 ntをもつ構築物(pTNT-264CAT)が最もCAT遺伝子を活性化した。pTNT-4000CAT〜pTNT-600CATは、pTNT-264CATの領域を含んでいるにもかかわらず、pTNT-264CATほどCAT遺伝子を活性化しなかった。これは、上流域に遺伝子発現を抑える領域があるためかもしれない。pCAT-BasicはCAT遺伝子の上流に何も連結しなかったもの、pCAT-Controlは、どんな細胞でも下流の遺伝子を活性化するSV40のprompterを連結した構築物である。
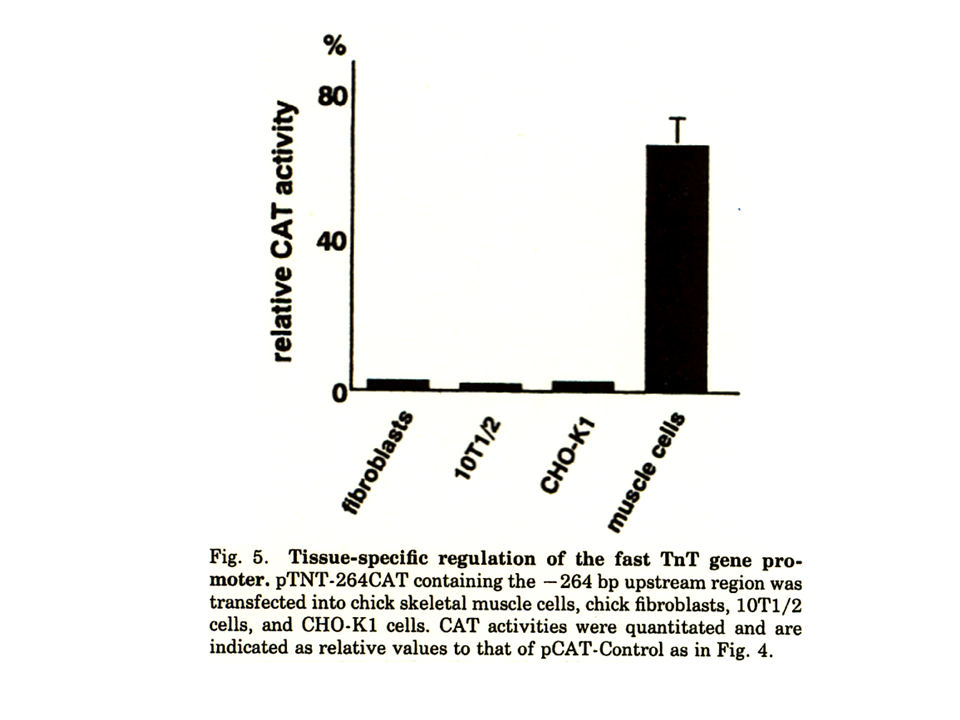
説明文の9
pTNT-264CATは筋細胞では、CAT遺伝子を活性化したが、繊維芽細胞や非筋培養細胞では、CAT遺伝子を活性化しなかった。
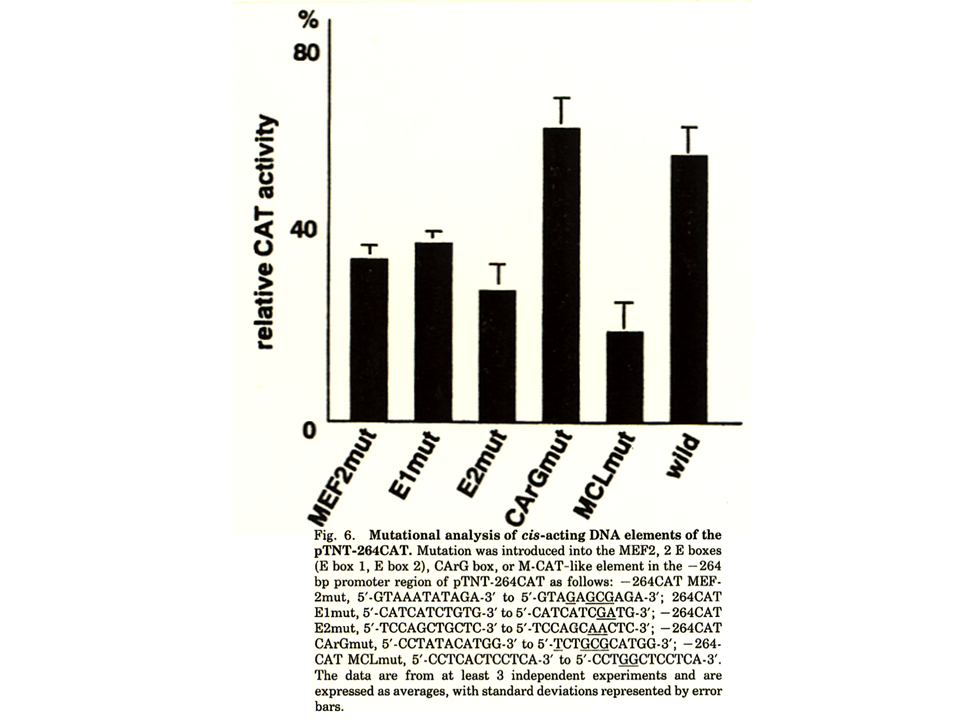
説明文の10
速筋型トロポニンT遺伝子の上流のcis-acting elementの候補の配列を変化させてmutantを作製した。CArG boxの塩基列を変化させた場合、何も変化がないもの(wild)より、CAT遺伝子をより活性化させたが、他の配列を変化させた場合は、CAT遺伝子の活性化がいくらか抑制された。特にM−CATと1塩基異なるM-CAT-like(MCL)では、CAT遺伝子の活性化が最も抑制された。
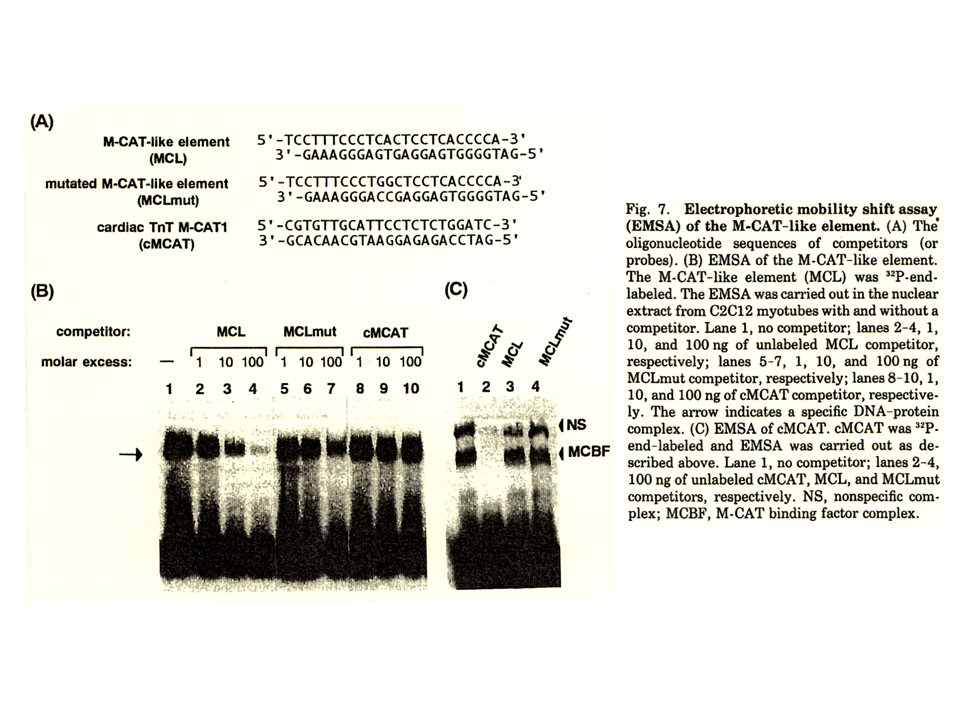
説明文の11
MCLが本当に速筋型トロポニンT遺伝子の発現調節にかかわるのか調べるために、EMSAを行った。MCLを含むプローブに核タンパク質が結合したことを示すバンドがみられた。ラベルされたプローブに対して、10倍量、100倍量のラベルされていないプローブを加えた場合、核蛋白質を奪い合うため、バンドが薄くなった。一方、MCLのmutantを含むラベルさえれていないプローブを10倍量、100倍量加えても、核タンパク質はMCL mutantを含むプローブに結合しないので、バンドは薄くならなかった。さらに、M-CATを含むプローブを10倍量、100倍量加えた場合にも、バンドは薄くならなかった。このことは、MCLと結合する核蛋白質があり、MCLが速筋型トロポニンT遺伝子の発現調節にかかわっている可能性が高いことを示している。また、MCLと1塩基しか異ならないM-CATには、この核蛋白質は結合しないことがわかった。

説明文の12
M-CATはニワトリの心筋型トロポニンT遺伝子の発現調節にかかわることが知られているが、1塩基しか異ならないMCLが速筋型トロポニンT遺伝子の発現調節にかかわっている可能性は高い。速筋型トロポニンT遺伝子のMCLのように、M-CATに類似した配列が遺伝子発現調節に関与している例がいくつか知られている。マウスのMyoD遺伝子の発現調節にかかわる配列は、MCLと全く同じ(CACTCCT)である。
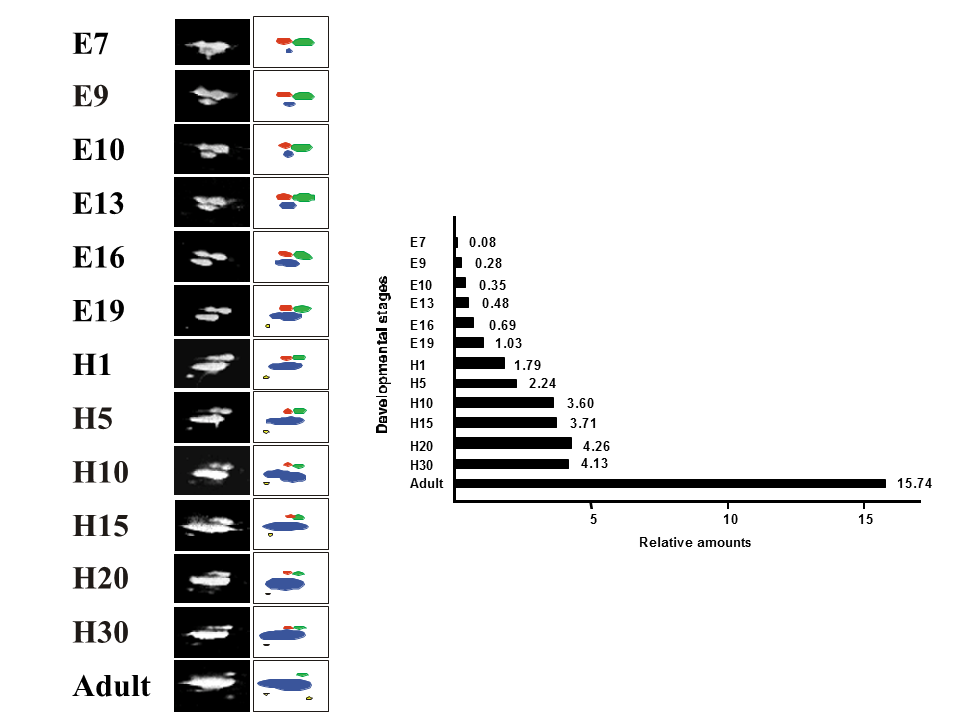
説明文の13
次に心筋型トロポニンTである。ニワトリ心筋型トロポニンT遺伝子は、選択的エクソンが1個あるので、2種類のアイソフォームを産生するが、翻訳後修飾(リン酸化)のために、2次元電気泳動パターン上で2つ以上のスポットがみられた。心臓での心筋型トロポニンTの発現を調べると、孵卵7日目には高分子量のスポット(赤色と緑色)の量が多く、低分子量のスポット(青色)の量は少なかったが、発生に伴って相対的に高分子量のスポットの量が減り、低分子量のスポットの量が増え、大人では低分子量のスポットが非常に多くなった。
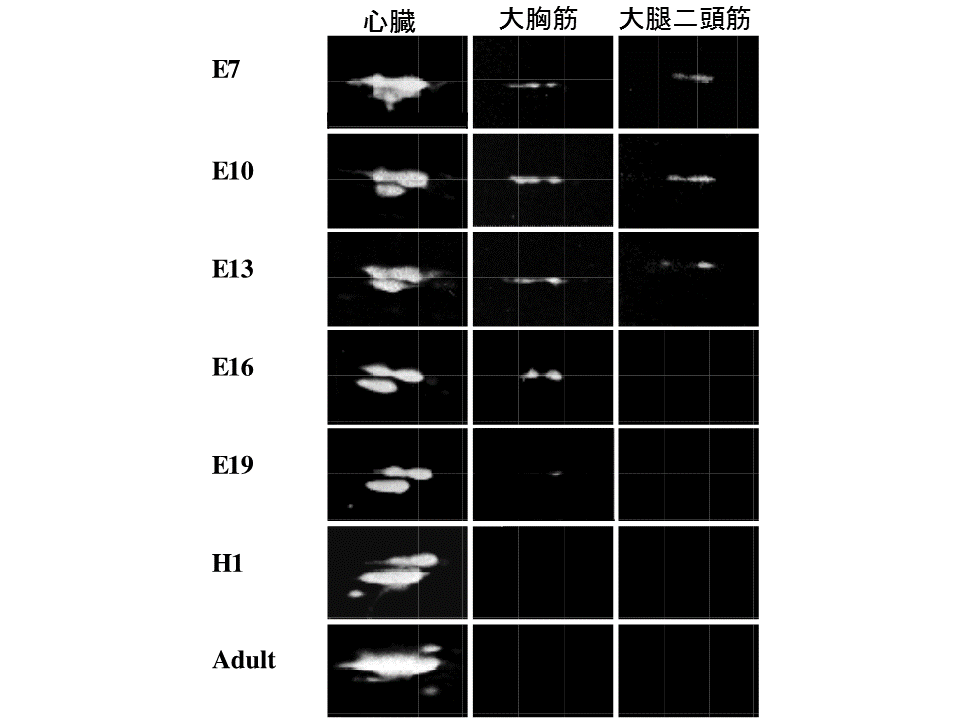
説明文の14
速筋の大胸筋や大腿二頭筋でも心筋型トロポニンTが孵化前後までみられた。発現している時期については、検出方法の違いや用いた個体の違いによって異なった。
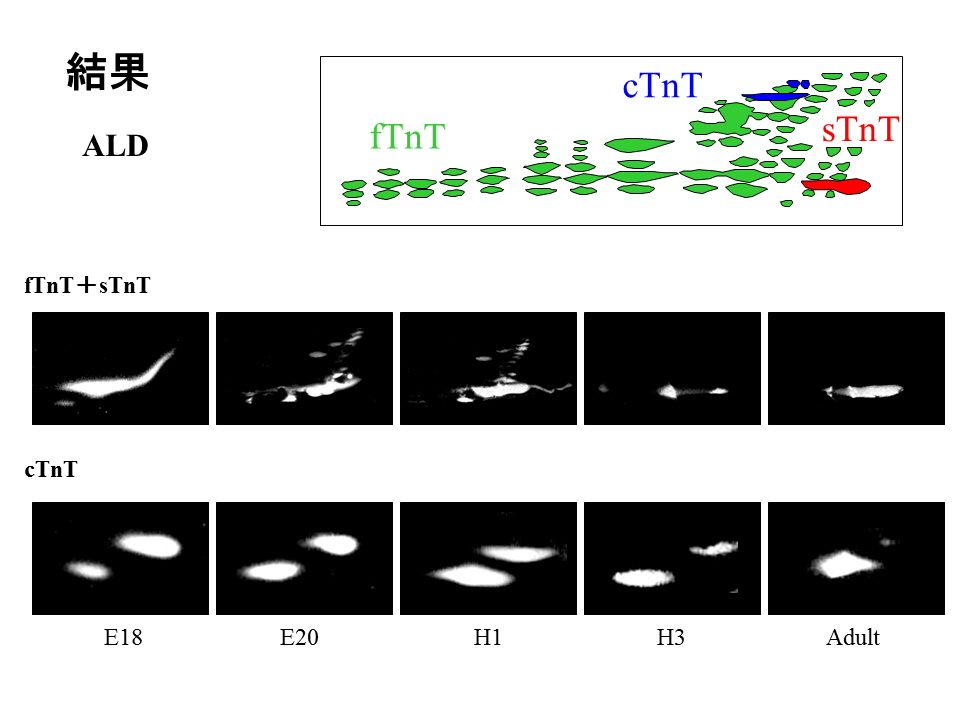
説明文の15
前広背筋では、遅筋型トロポニンT(sTnT)は発生を通してみられ、速筋型トロポニンT(fTnT)は孵化後3日目までみられた。心筋型トロポニンT(cTnT)は孵化前後までみられると考えられていたが、この実験では大人の前広背筋にもcTnTがみられ、調べた全発生段階を通して発現しているということがわかった。
説明文の16
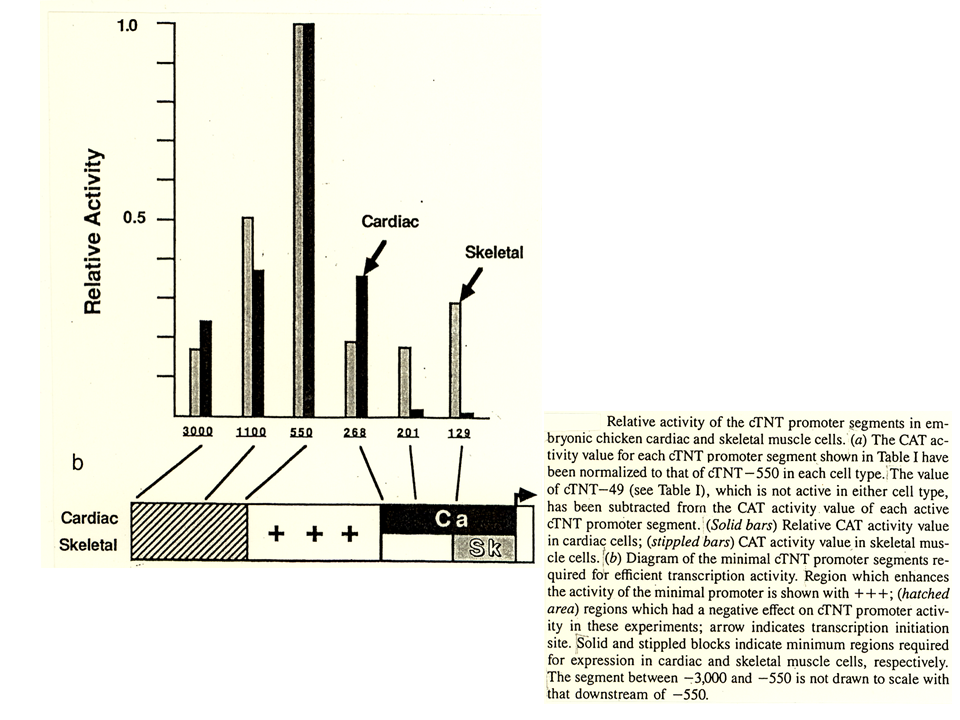
最後に心筋型トロポニンT遺伝子の発現調節に関する研究を紹介しよう。心筋型トロポニンT遺伝子の上流域をCAT遺伝子に結合してCAT assayを行った結果、心筋から得た培養細胞を用いても、骨格筋から得た培養細胞を用いても、上流550 ntを連結した場合に最もCAT遺伝子を活性化した。しかし、上流201 ntを連結した場合には、心筋培養細部でCAT 遺伝子の活性が極端に落ちてしまう。
説明文の17
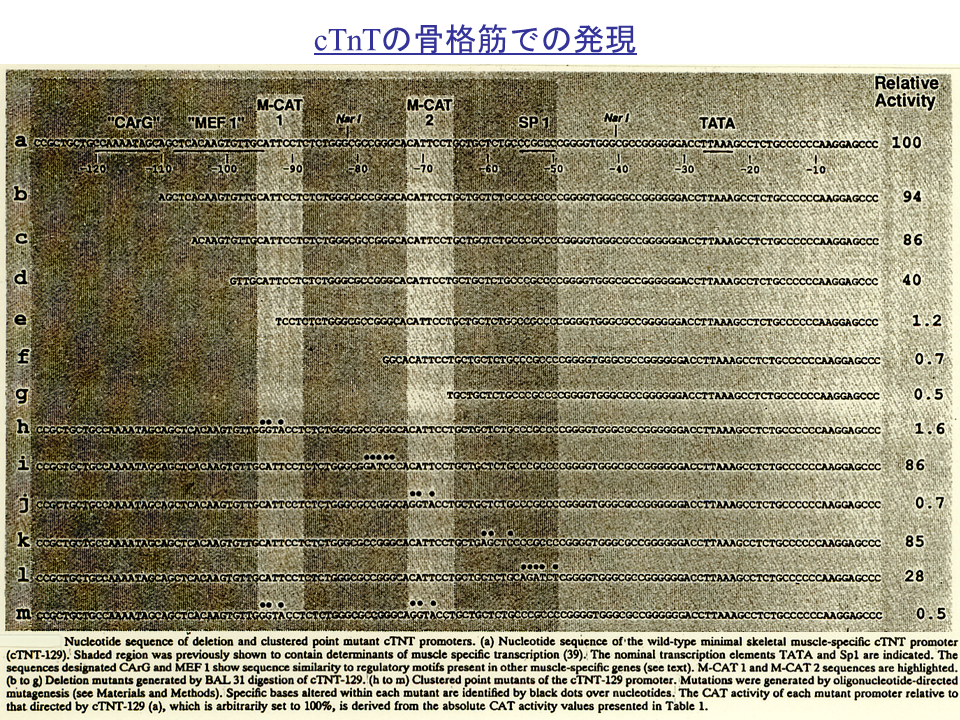
遺伝子の上流129 ntまでに、M-CATが2か所にあるなど、遺伝子発現調節にかかわる可能性のある配列が複数ある。しかし、心筋の決定や分化には、MyoD、Myf5、myogeninはかかわらないので、それらの蛋白質の結合するE-boxはみられない。上流側のM-CATの中央辺りまで上流域を削ると、CAT遺伝子の活性化は極端に抑えられる。また、上流側のM-CATの塩基配列を変えても、下流側のM-CATの塩基配列を変えても、CAT遺伝子の活性化は極端に抑えられる。M-CAT以外の配列を削っても、配列を変えても、M-CATほどCAT遺伝子の活性化は抑制されない。したがって、2つのM-CATのどちらも心筋型トロポニンT遺伝子の発現調節には重要であることがわかる。
説明文の18
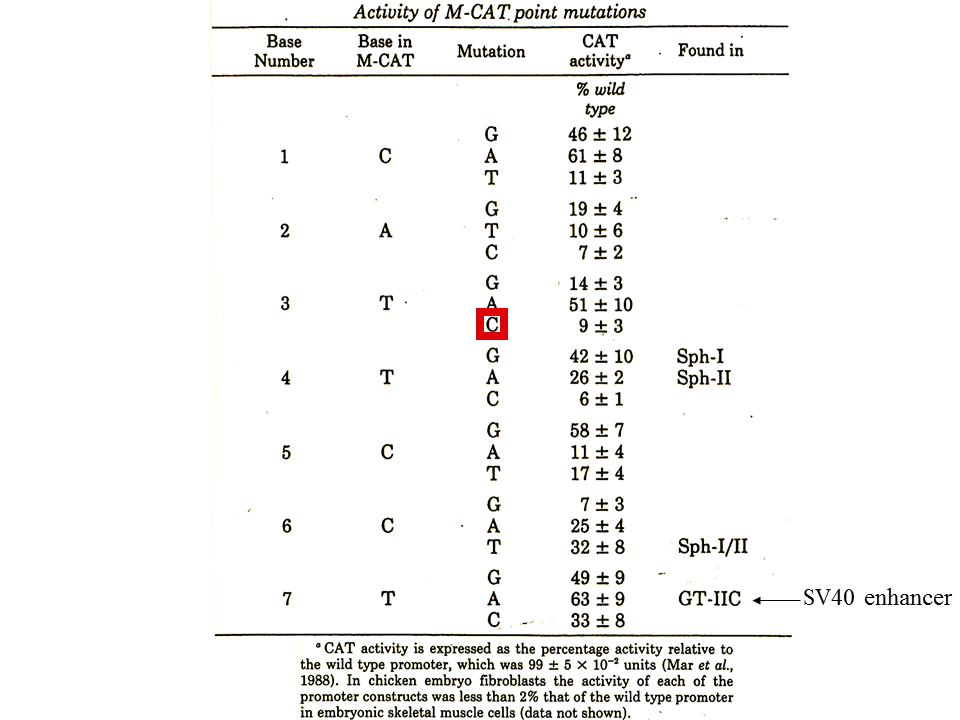
M-CAT(CATTCCT)の塩基の一つ一つを変化させてCAT assayを行うと、どの変化によってもCAT遺伝子の活性化は抑えられる(6~63%)。注目すべきなのは、CATTCCTを速筋型トロポニンンT遺伝子の発現調節に重要であると考えられるMCLのCACTCCTに変化させた場合(赤色の四角で囲んだ)、CAT遺伝子の活性化は9%に抑えられる。したがって、M-CATとMCLは1塩基異なるだけであるが、機能は全く異なるのである。
説明文の19
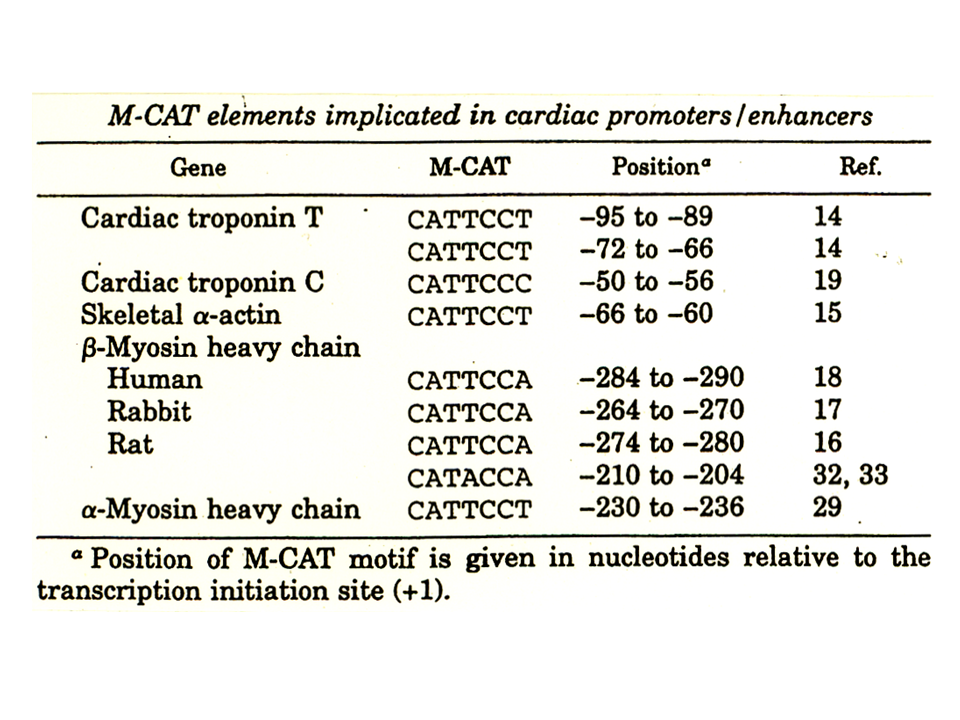
M-CATは他の様々な遺伝子の発現調節にかかわっている。また、M-CATとは1あるいは2塩基異なる配列も同様に発現調節にかかわっている。

説明文の20
心筋型トロポニンンT遺伝子の発現調節はさらに複雑である。M-CATのどちらかの配列を変化させるとCAT遺伝子の活性化は極端に抑えられるが(kとl)、M-CATだけでは不十分である。上流(201から269 まで)の配列を削るとCAT遺伝子の活性化は極端に抑えられる(bとc)。この領域は遺伝子の発現を活性化するエンハンサーであると思われる。エンハンサーは配列を逆転させても機能する(d~g)。上流(201から269 まで)のうち、247から269までの配列はエンハンサーの機能に関係がないらしい(h)。201~215の配列だけでは、CAT遺伝子の活性化は極端に抑えられる(iとj)。これは後述するが、この部分には核蛋白質が結合できないためである。
説明文の21
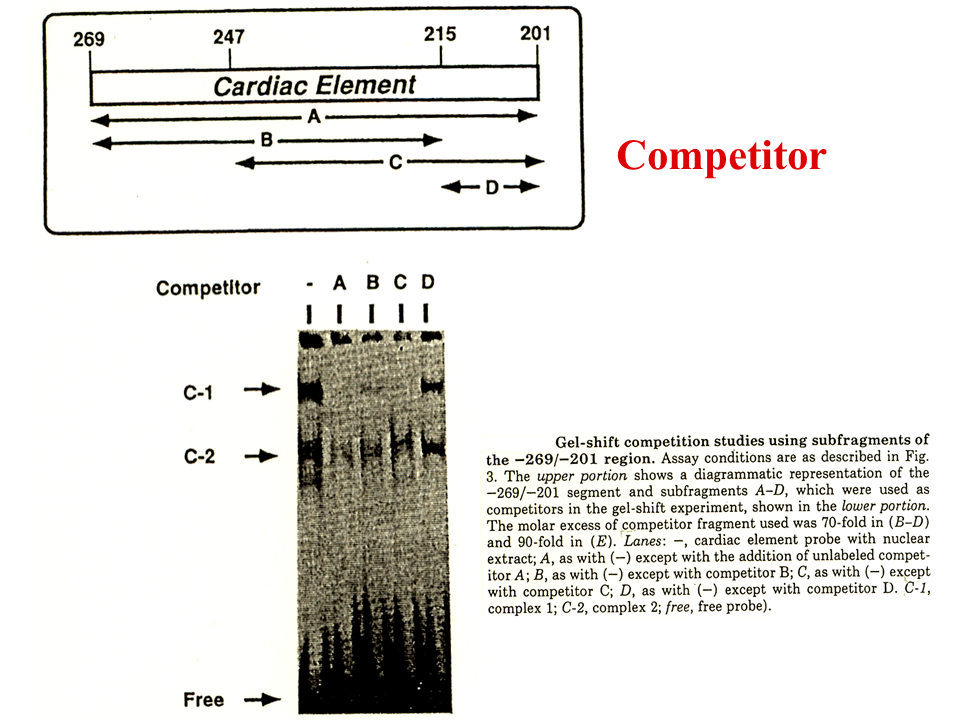
上流(201から269 まで)の配列には核蛋白質が結合する(-)。この領域を切断し、各々を過剰に加えた場合、競合して、A、B、CではC-1の部分の核蛋白質の結合がみられなくなる。Dは核蛋白質の結合に影響しない。したがって、215~247に核蛋白質は結合することがわかる。

説明文の22
まとめると201~269の配列の内、247~269は何の機能もない。201~247はエンハンサーとして機能するためには必要であり、特に215~247は核蛋白質と結合する。
説明文の23
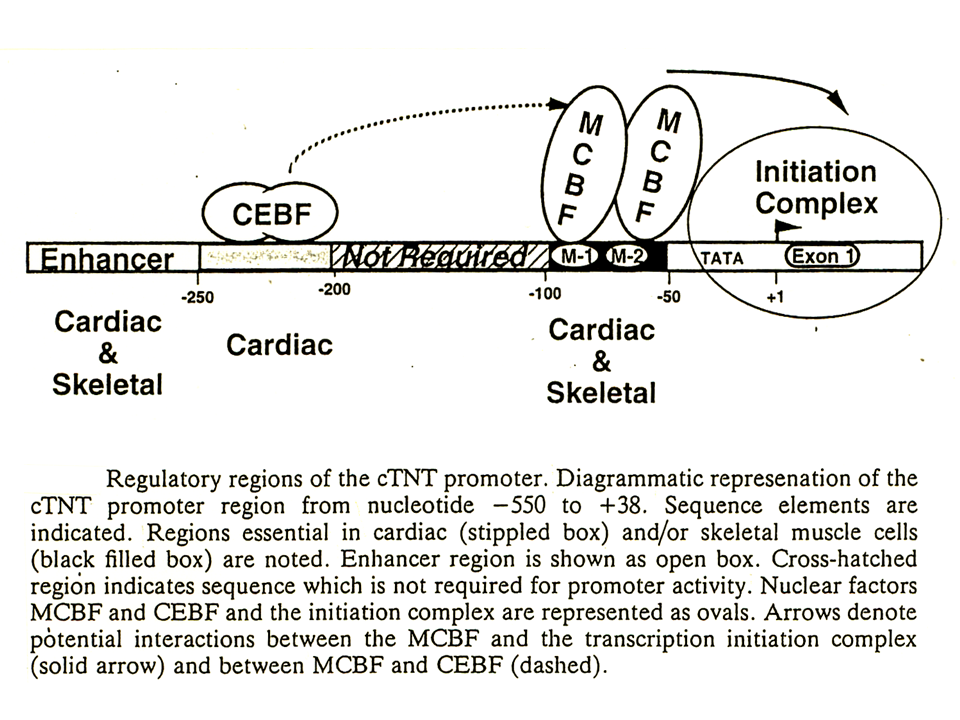
心筋型トロポニンT遺伝子の発現調節の全体像をまとめると、2つのM-CATは心筋型トロポニンT遺伝子の発現調節に重要である。ここには核蛋白質MCBFが結合する。上流の215~247にも各蛋白質CEBFが結合し、MCBFに影響を与えて心筋型トロポニンT遺伝子の発現を活性化する。
説明文の24
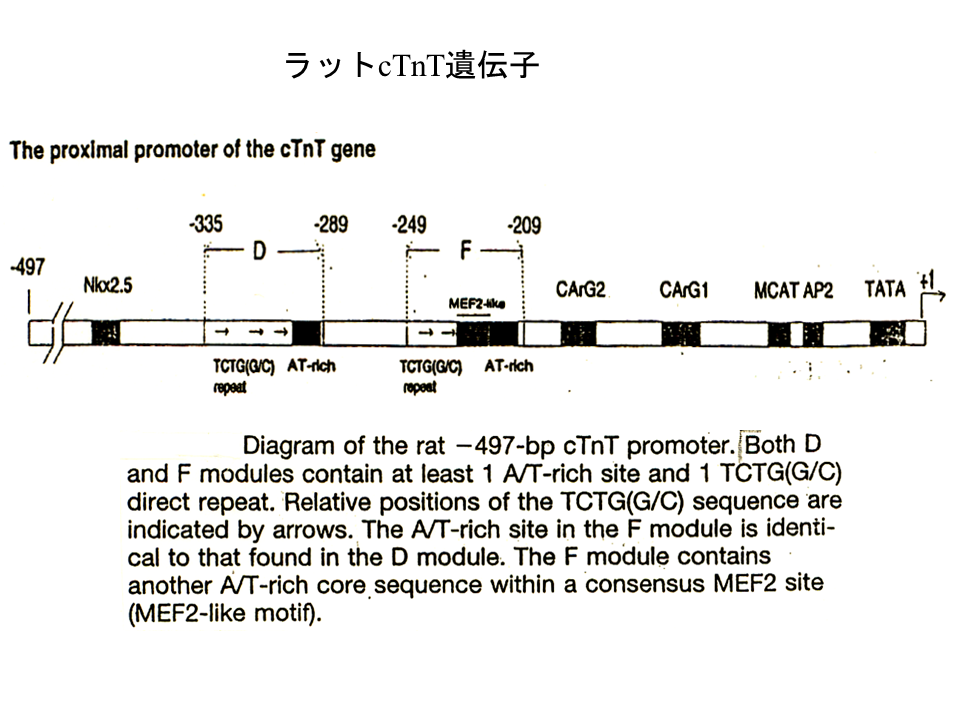
それでは他の脊椎動物の心筋型トロポニンT遺伝子でも、ニワトリの遺伝子と同様に発現調節が行われるのであろうか?ラットの心筋型トロポニンT遺伝子の上流にも、遺伝子の発現調節にかかわる可能性のある配列が複数みられる。M-CATも存在する。
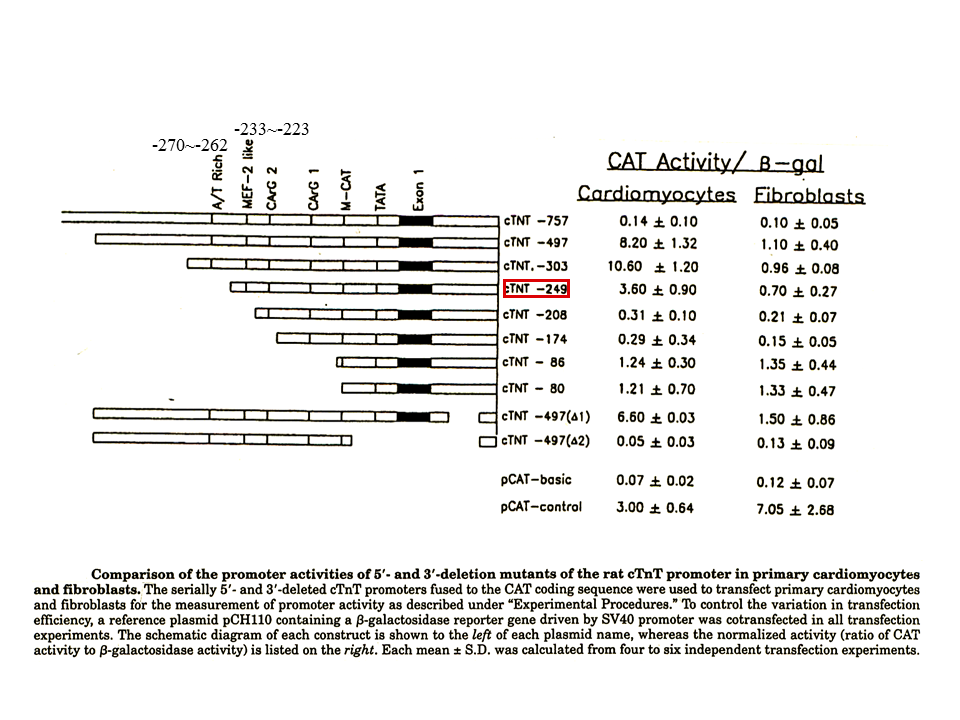
説明文の25
心筋細胞(Cardiomyocyte)では、上流部を徐々に削っていった場合、上流249 ntを含む断片まで削っても、CAT 遺伝子の活性化が十分みられる(cTNT-249)。しかし、208 ntを含む断片まで削ってしまうと、CAT 遺伝子の活性化は極端に抑えられる。すなわち、208から249までに重要な配列が存在することを示している。心筋細胞に比べ繊維芽細胞(Fibroblast)では、CAT 遺伝子の活性化は低い。

説明文の26
そこにはM-CATではなく、MEF-2 likeが存在した。MEF-2に共通した配列はCAT(A/G) 4TAGであるが、MEF-2 likeの配列はTTTAAATAGであった。すなわちニワトリ心筋型トロポニンT遺伝子の場合と異なり、M-CATは関係ないのである。したがって、同じ心筋型トロポニンT遺伝子であっても、その発現調節は動物種によって異なるのである。
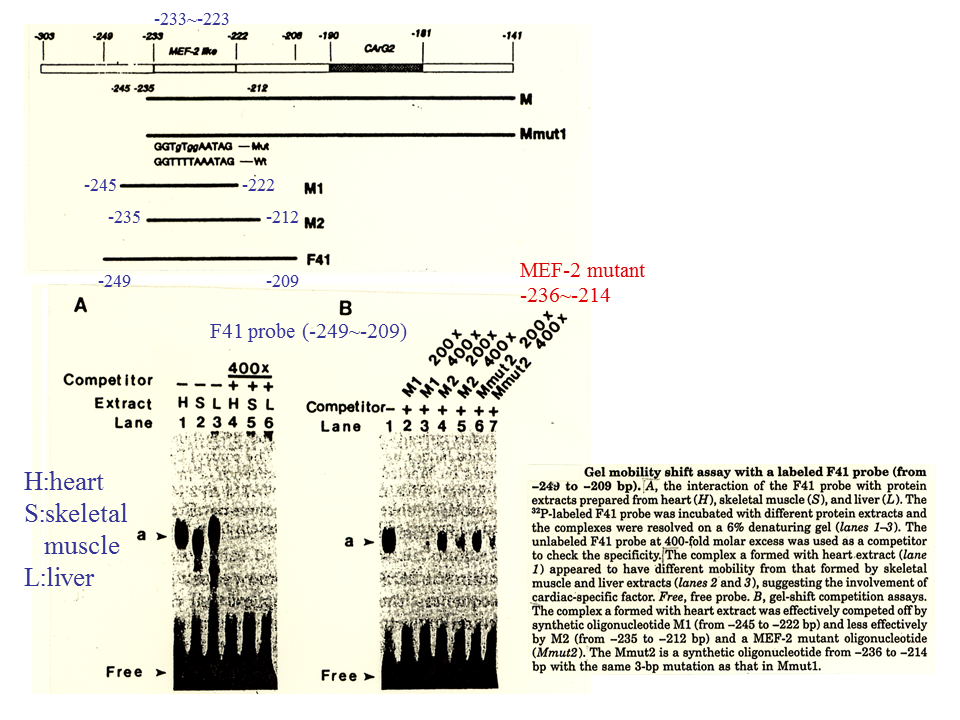
説明文の27
ラット心筋型トロポニンT遺伝子の上流域の209~249を含む断片F41は、心臓、骨格筋、肝臓の核蛋白質と結合する。ただし、結合する核蛋白質の移動度は心筋のものと骨格筋、肝臓のものとでは異なる。ラベルしないF41を過剰量加えると、競合してバンドがみられなくなる。また、222~245を含む断片M1を過剰量加えても、競合してバンドがみられなくなる。したがって、核蛋白質は222~245に結合する。212~235を含む断片M2を過剰量加えた場合、いくらか競合してバンドが薄くなる。MEF-2 likeは223~233に存在するのが、核蛋白質はこれよりもやや上流に結合するようにもみえる。いずれにしても、鳥類と哺乳類では、心筋型トロポニンT遺伝子の調節機構が異なるようである。
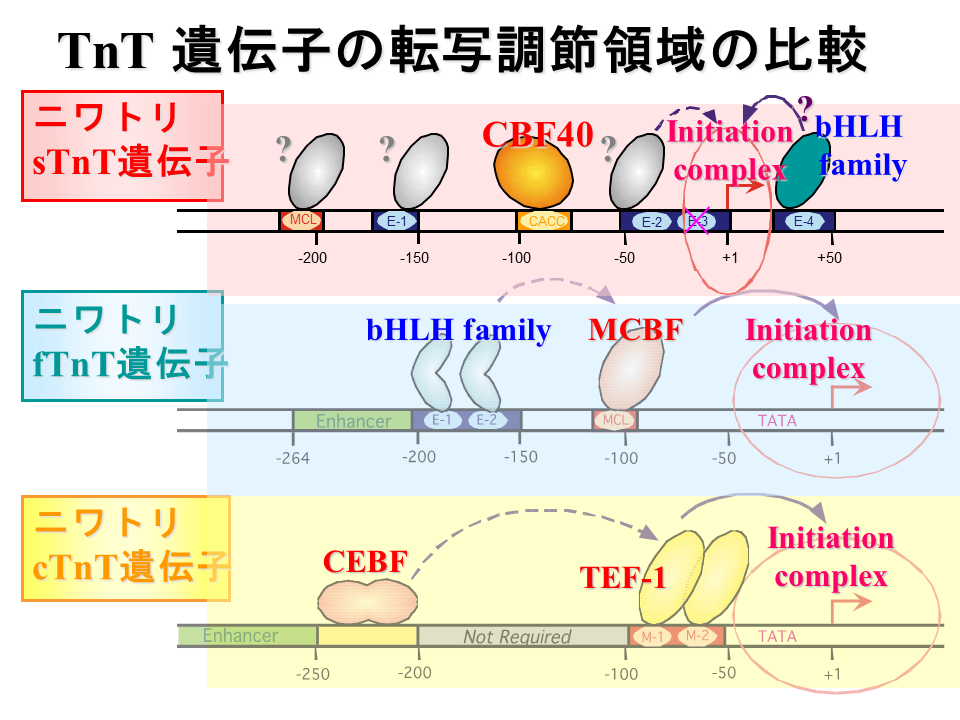
説明文の28
ニワトリトロポニンT遺伝子の発現調節において、遅筋型遺伝子ではE-box4がcis-acting elementとして働くと思われ、CACC-boxも何らかの影響を与えるかもしれない。E-boxはMyoD、Myf5、myogeninの結合に関与する可能性がある。速筋型遺伝子では同様にE-boxがMyoD、Myf5、myogeninの結合に関与し、MCL(M-CAT-like)がcis-acting elementとして働く。心筋型遺伝子では、M-CATがcis-acting elementとして働き、上流の領域がそれに影響を与える。心筋型遺伝子ではM-CATが、速筋型遺伝子ではそれとは1塩基異なるMCLが重要であるが、遅筋型遺伝子ではM-CATと2塩基異なるMCLは何の機能ももたないと思われる。骨格筋の細分化あるいは心筋の分化は、つきつめれば例えばトロポニンT遺伝子では、これらのcis-acting elementに結合するtrans-acting factorの違いによって引き起こされる。では、trans-acting factorの違いはどのようにして生み出されるのか?これはまたさらに上流の遺伝子によって制御されるのかもしれない。この階層的な構造をどこまでさかのぼればよいのか、今後の課題である。