説明文の1
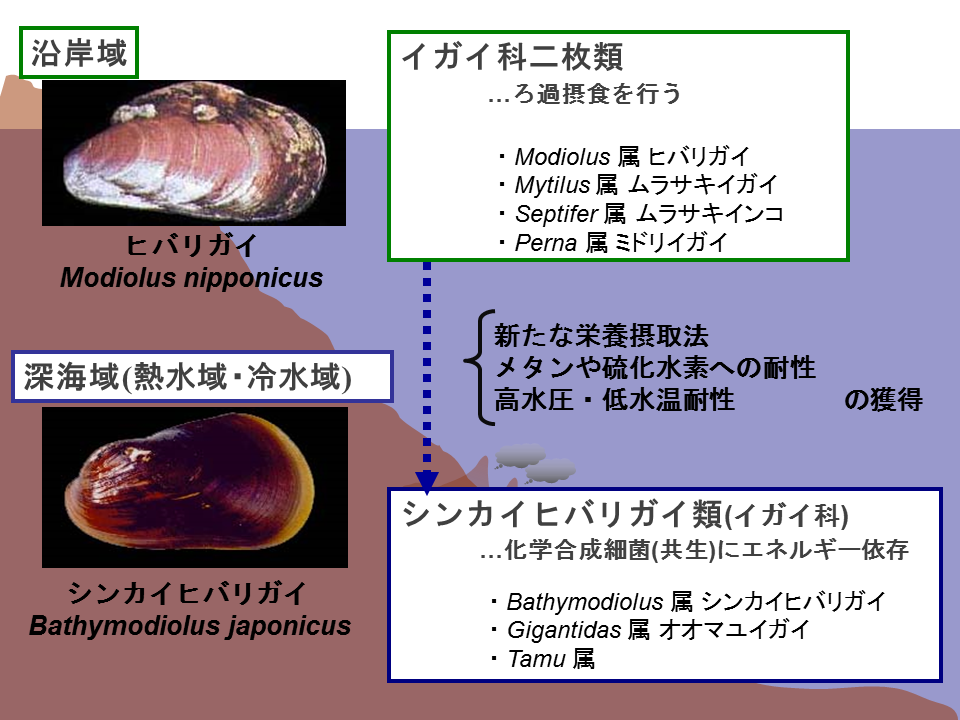
イガイ類は沿岸域に卓越しており、浅海域から深海域に進出したと考えられる。浅海域のイガイ類は共生細菌をもたず、ろ過摂食のみによって栄養を得ているため、深海域への進出のためには新たな栄養摂取法を獲得する必要がある。また、メタンや硫化水素への耐性、高水圧や低水温への耐性なども獲得する必要がある。
説明文の2
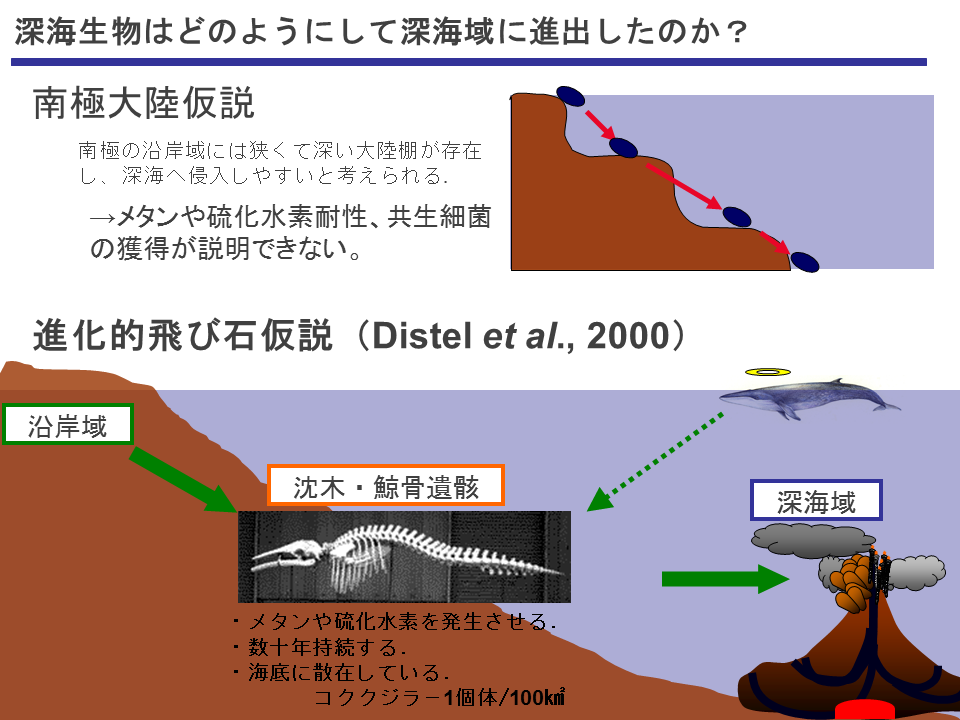
イガイ類の深海域への進出を説明する仮説として、南極大陸の特徴的な狭くて深い大陸棚を利用したとする「南極大陸仮説」と海底に散在する沈木や鯨骨遺骸を足がかりにしたとする「進化的飛び石仮説(進化的ステッピングストーン仮説)」がある。
説明文の3
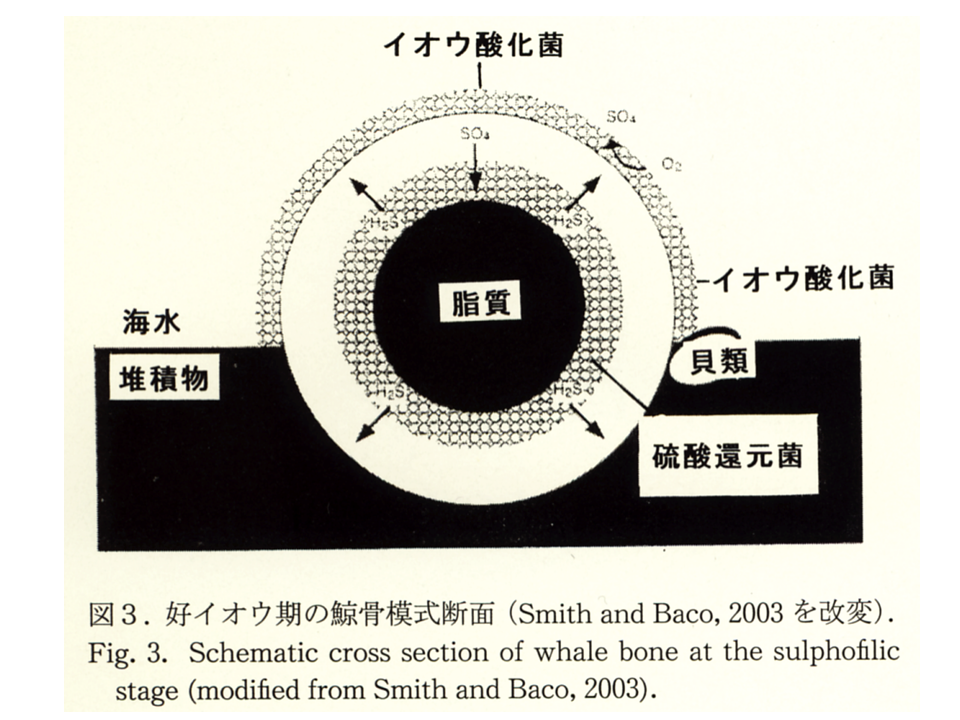
沈木や鯨骨遺骸は嫌気的に分解され、数十年間メタンや硫化水素を発生させるため、化学合成細菌と共生関係を獲得する場になりうる。また、イガイ類が足糸を使って付着する基質にもなる。鯨の骨の中には鯨油が収容されている。
説明文の4
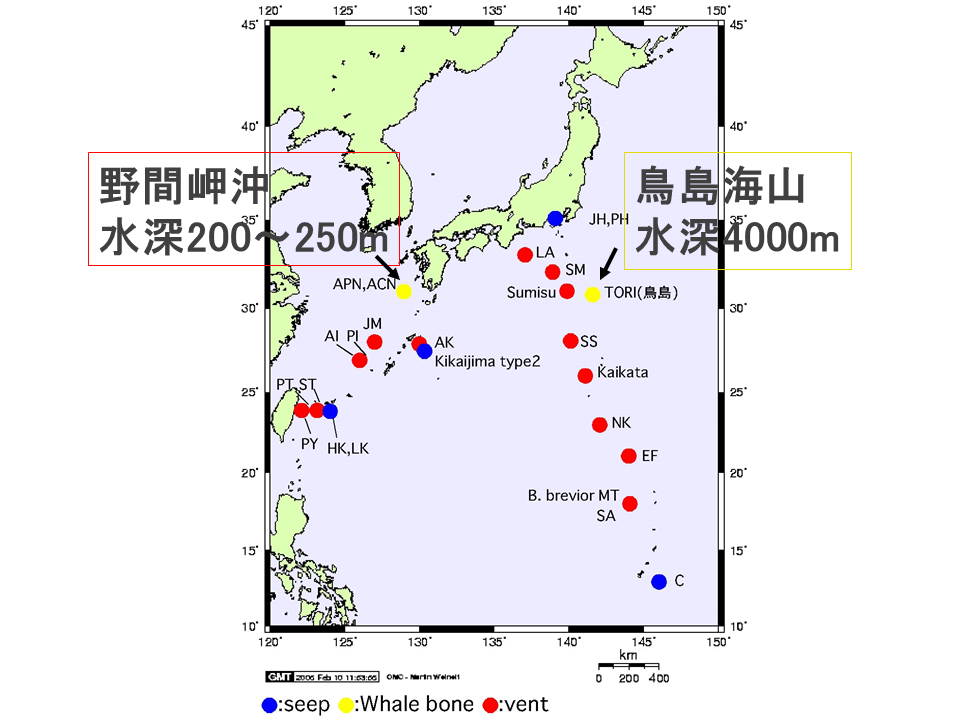
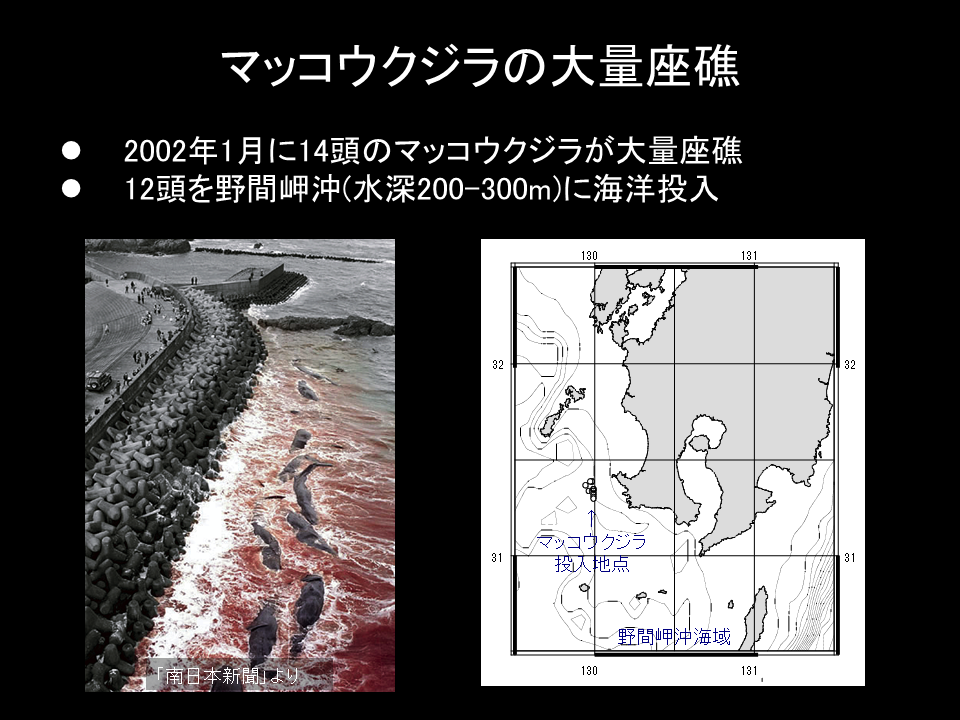
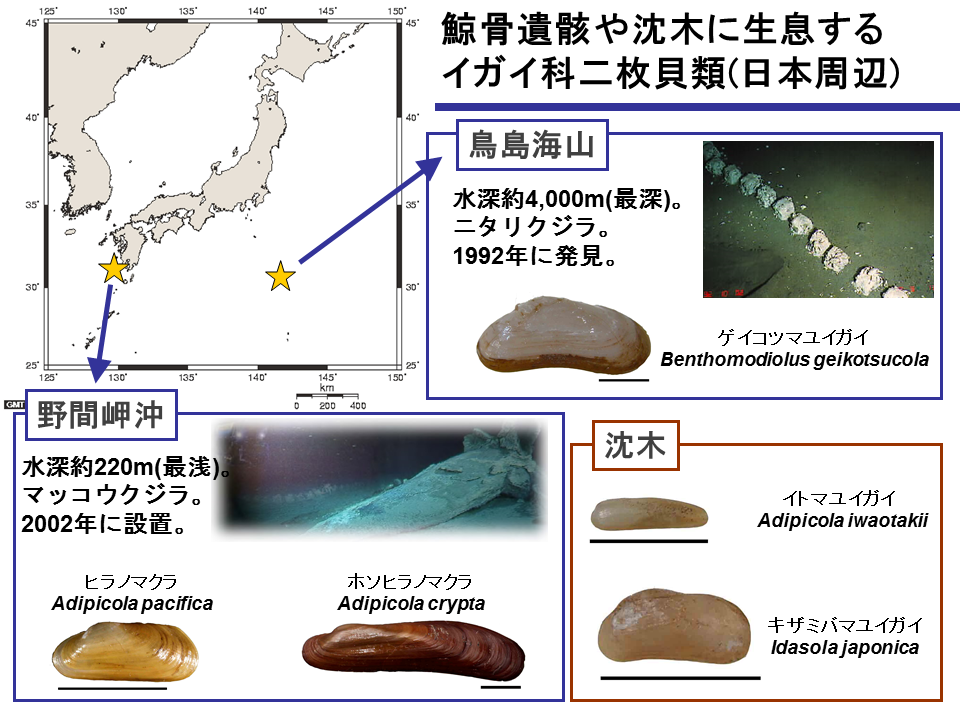
日本周辺海域で発見されている自然死した鯨遺骸は、鳥島海山(水深約4000m)のニタリクジラのものだけである。座礁したマッコウクジラの遺骸を野間岬沖(水深約250m)に人為的に設置した。また、相模湾(水深約1000m)にもマッコウクジラなどの遺骸を人為的に設置した。
説明文の5
2002年に座礁したマッコウクジラを野間岬沖に設置した。

説明文の6
設置したマッコウクジラ遺骸の変化。鯨骨は急速に崩壊し、堆積物に覆われていった。

説明文の7
無人潜水船ドルフィン3Kで、野間岬沖の人工的に設置した鯨遺骸から鯨骨を回収した。母船上に引き上げられた鯨骨に付着したイガイ類(ヒラノマクラAdipicola pacifica)。

説明文の8
野間岬沖の鯨骨からヒラノマクラとホソヒラノマクラが採集された、ヒラノマクラは海水中に露出した鯨骨に多く、ホソヒラノマクラは堆積物中に埋没した鯨骨に多かった。硫化水素の濃度に応じて棲み分けが起こっているのだろうか。
説明文の9
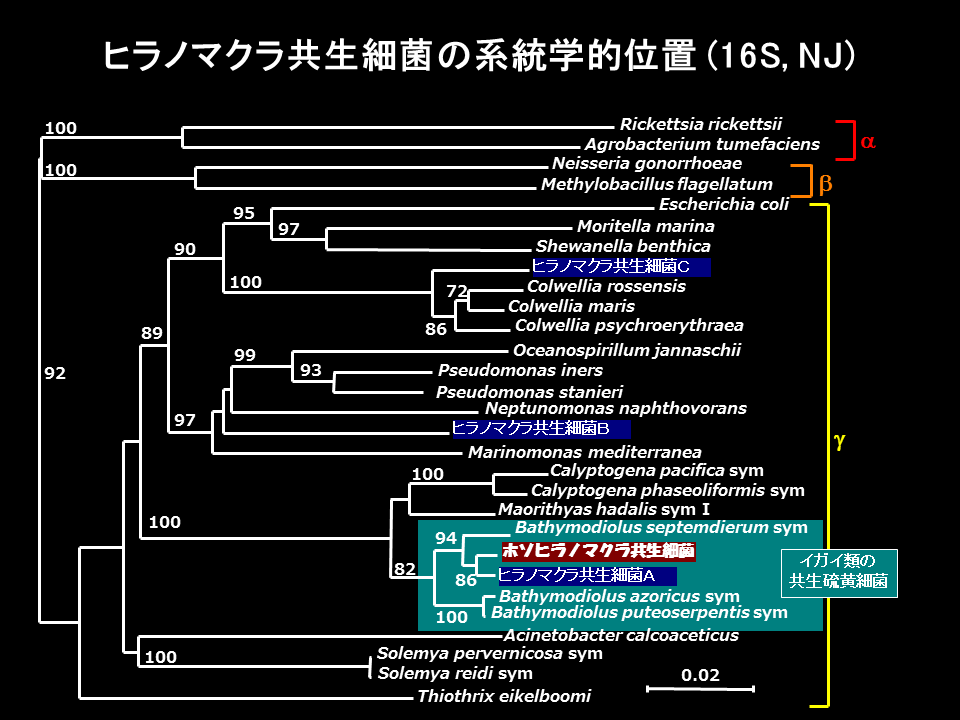
ヒラノマクラの鰓には数種の共生細菌がみられたが、その一つはシンカイヒバリガイ類の硫黄細菌に近縁であった。また、ホソヒラノマクラの共生細菌もシンカイヒバリガイ類の硫黄細菌に近縁であった。
説明文の10
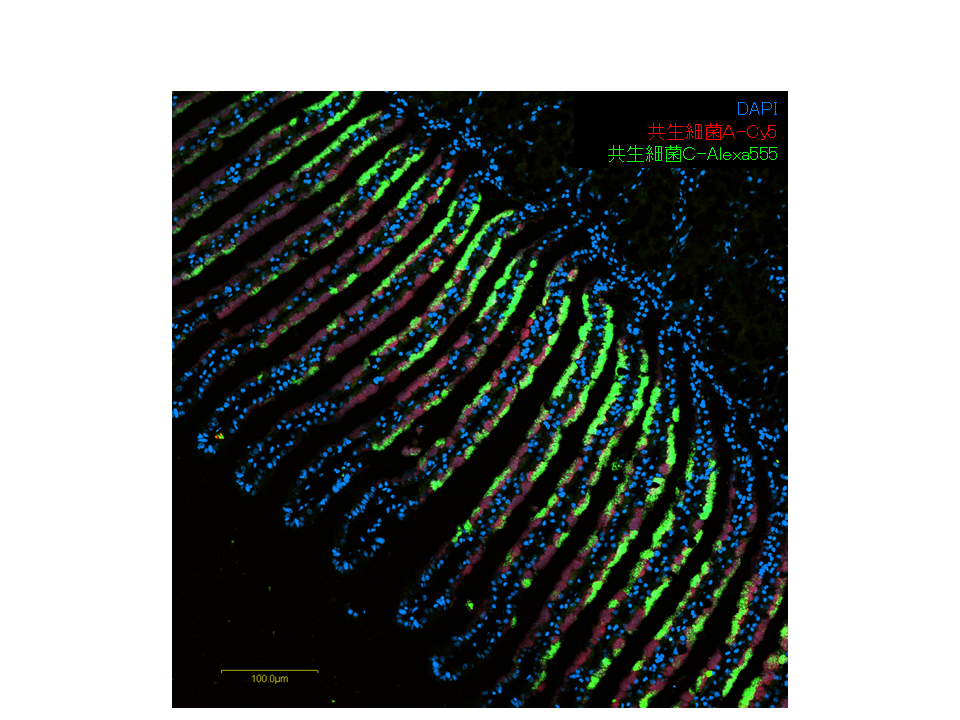
共生する細菌の間で鰓での局在が異なる。赤と緑は2種類の細菌を示している。共生細菌Aは硫黄細菌。青は核。

説明文の11
日本周辺海域で見つかっている唯一の自然死した鯨の遺骸を調査するため、「しんかい6500」の母船である「よこすか」に乗船した。

説明文の12
「深海2000」と同様に、船長、船長補佐、研究者の3名が「しんかい6500」に乗り込んだ
説明文の13
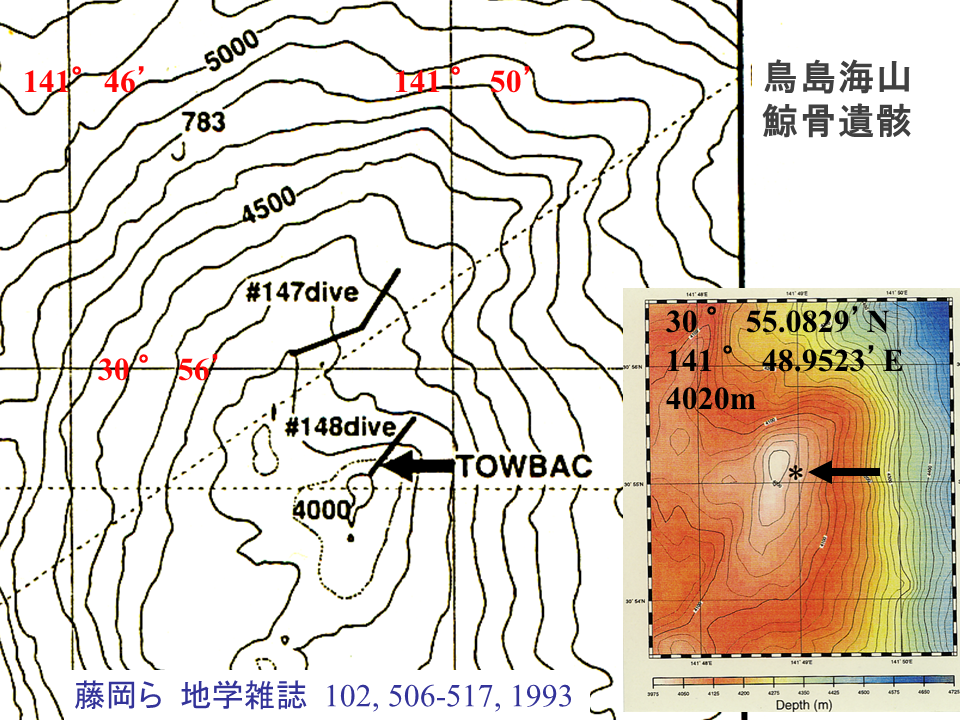
鯨遺骸は論文に示されているものと異なる位置に存在した。GPS装置の精度の違いによるらしい。実は着底する位置が計画通りではなかったが、これがむしろ幸いして鯨遺骸をすぐにみつけることができた。野間岬沖に人為的に設置した鯨遺骸の変化を考えると、発見してから10以上経過している鳥島海山の鯨遺骸が見つかるのか、あったとして鯨油が枯渇せずにまだ生物を養っているのか、不安は大きかった。
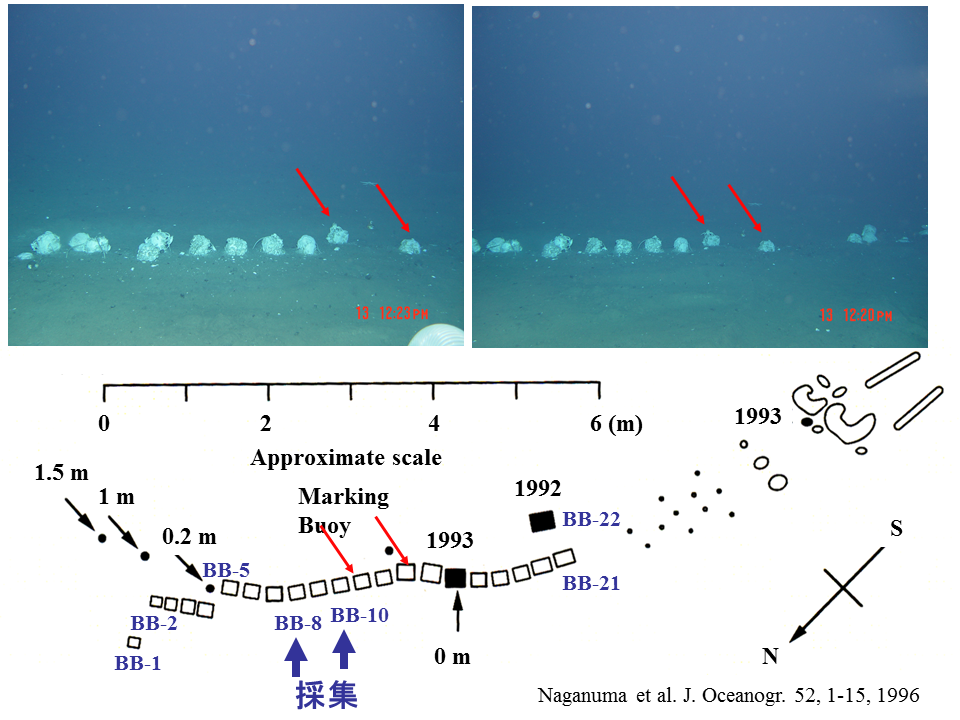
説明文の14
鳥島海山で1992年に発見された鯨遺骸は2005年の潜航でも、大きく崩壊することもなく、また堆積物に埋没することもなく存在した(上の写真)。野間岬沖(水深約250m)の鯨遺骸は短い期間で崩壊し、堆積物におおわれた。おそらく水深が深いため(水深約4000m)、発見から10年以上たっても大きな変化がなかったと思われる。赤の矢印は1993年に発表された骨の配置図上(下の図)で示したものと同じ骨を示す。

説明文の15
顎の骨の崩壊は脊椎骨よりいくらか速いようにみえる(左上と右下の写真)。赤の矢印は1993年に発表された骨の配置図上(右上の図)で示したものと同じ骨を示す。

説明文の16
脊椎骨BB-8とBB-10を採集した。骨の上に生物がいることがわかる。

説明文の17
母船上に引き上げられた鯨骨に付着したイガイ類。 2005 年の調査を基にこのイガイ類はゲイコツマユイガイ(Benthomodiolus geikotsucola )として新種記載された。
説明文の18
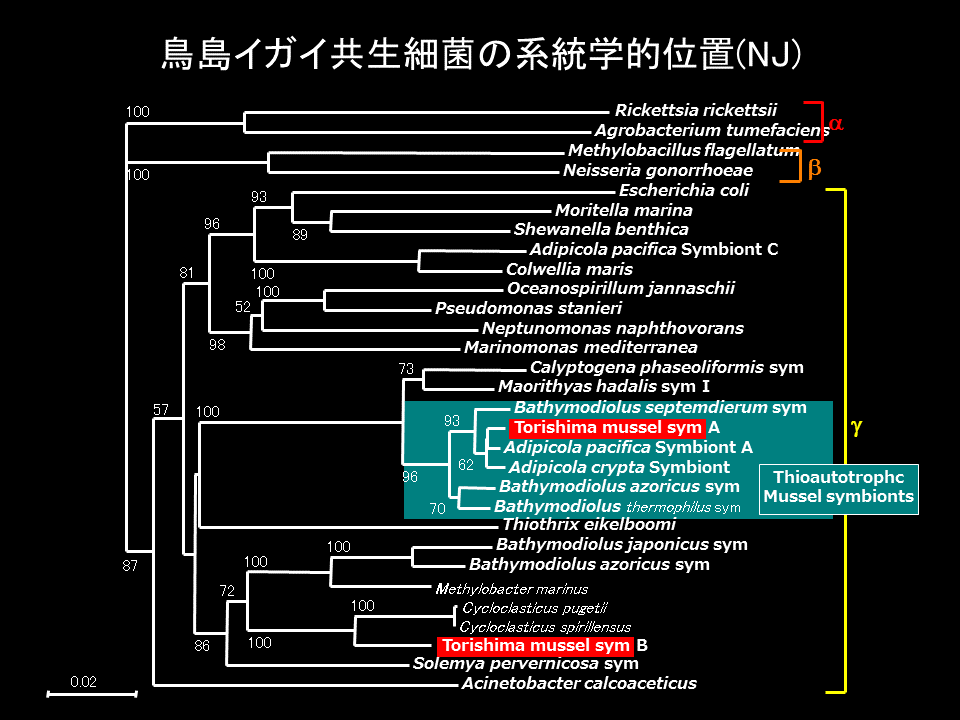
ゲイコツマユイガイの鰓にも共生細菌が存在し、それらはシンカイヒバリガイ類の硫黄細菌に近縁であった。
説明文19
野間岬沖や鳥島海山のイガイ類に、他の鯨遺骸や沈木からのイガイ類を加え、遺伝学的解析を行った。
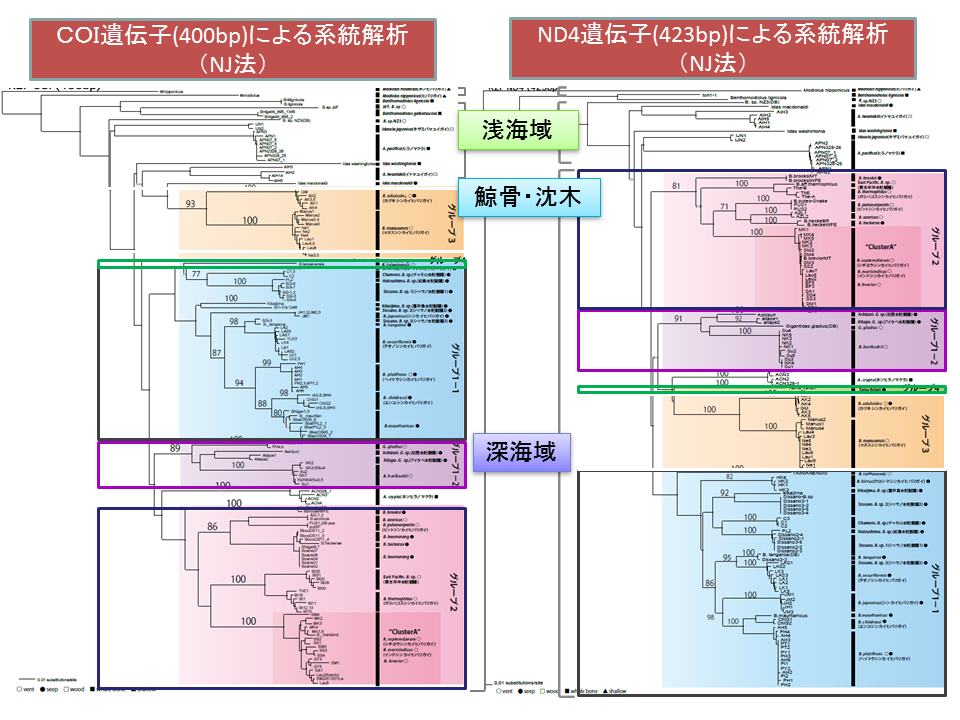
説明文の20
ミトコンドリアのCOI遺伝子とND4遺伝子の各々の塩基配列を基に構築した系統樹。どちらでも、深海域(熱水域・湧水域)に生息するシンカイヒバリガイ類の外群に鯨骨遺骸・枕木のイガイ類(例外はホソヒラノマクラAdipicola crypta)、さらにその外側に浅海域のイガイ類がみられ、「進化的飛び石仮説」を支持している。鯨骨遺骸・枕木のイガイ類の中にはシンカイヒバリガイ類とは独立に熱水域・湧水域に進出したものがいる。グループ1-1、1-2、2、3、4の関係は両樹形図で異なった。
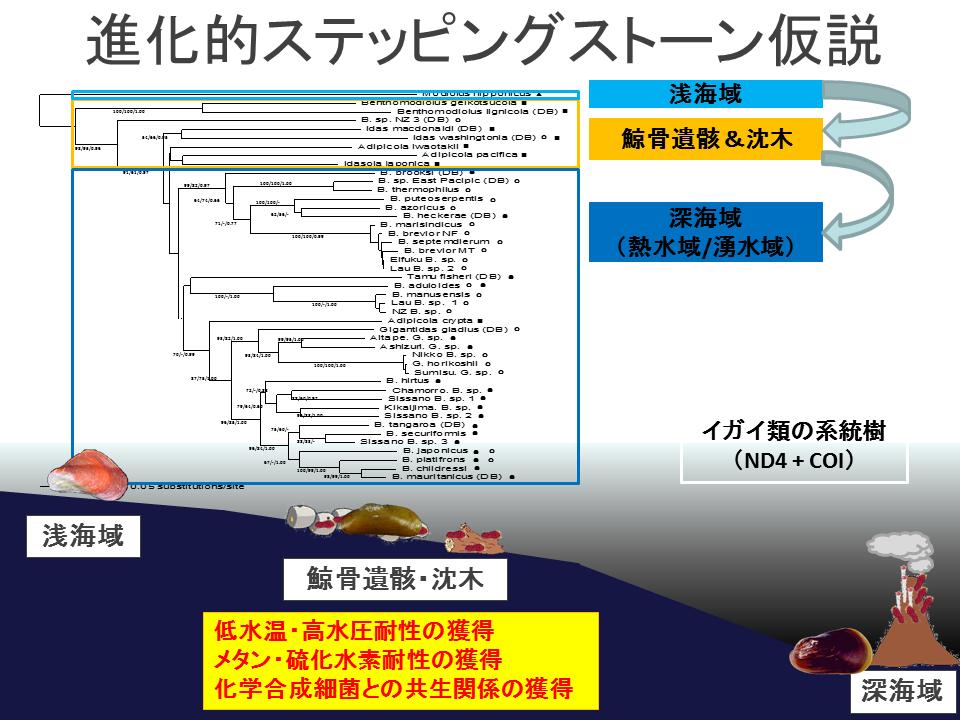
説明文の21
イガイ類は 、浅海域 、鯨骨遺骸・沈木、 深海の熱水域、湧水域という順番に分岐していく。 この結果はイガイ類において、 進化的ステッピングストーン仮説が支持されることを示している。すなわち、現生の深海生イガイ類は、過去に鯨骨遺骸を中継して進化してきたと考えられる。
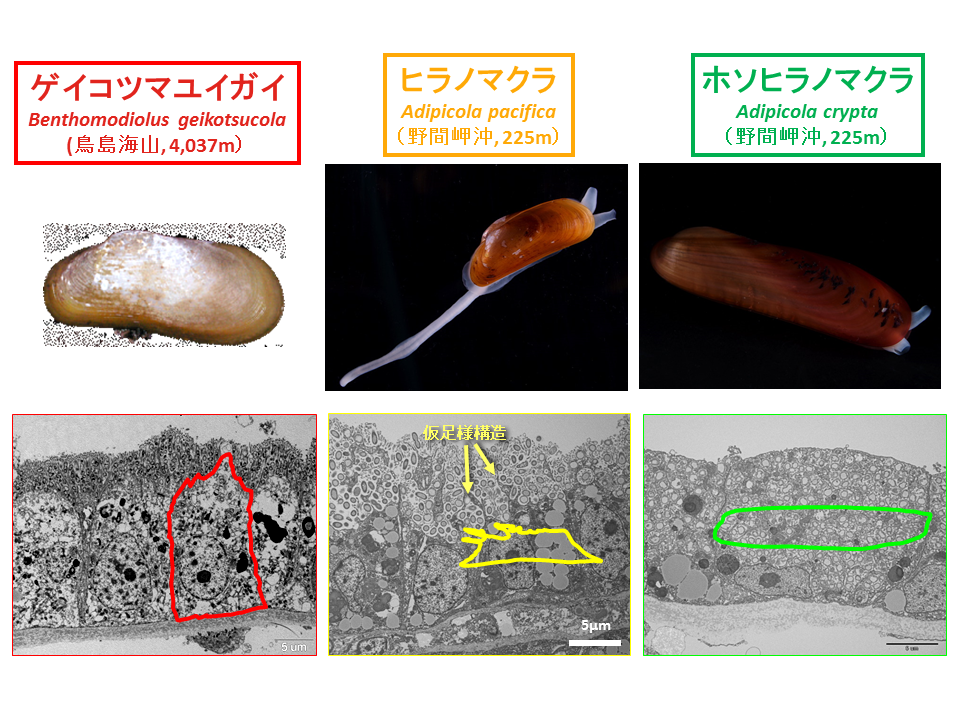
説明文の22
鯨骨に付着していたイガイ類の鰓上皮細胞を電子顕微鏡観察すると、ゲイコツマユイガイはとヒラノマクラは細胞外に、ホソヒラノマクラは細胞内に化学合成細菌を共生させていた。ゲイコツマユイガイは微絨毛に細菌を付着させ、ヒラノマクラは細胞膜が突き出して細菌を囲みこんでいるようにみえる。
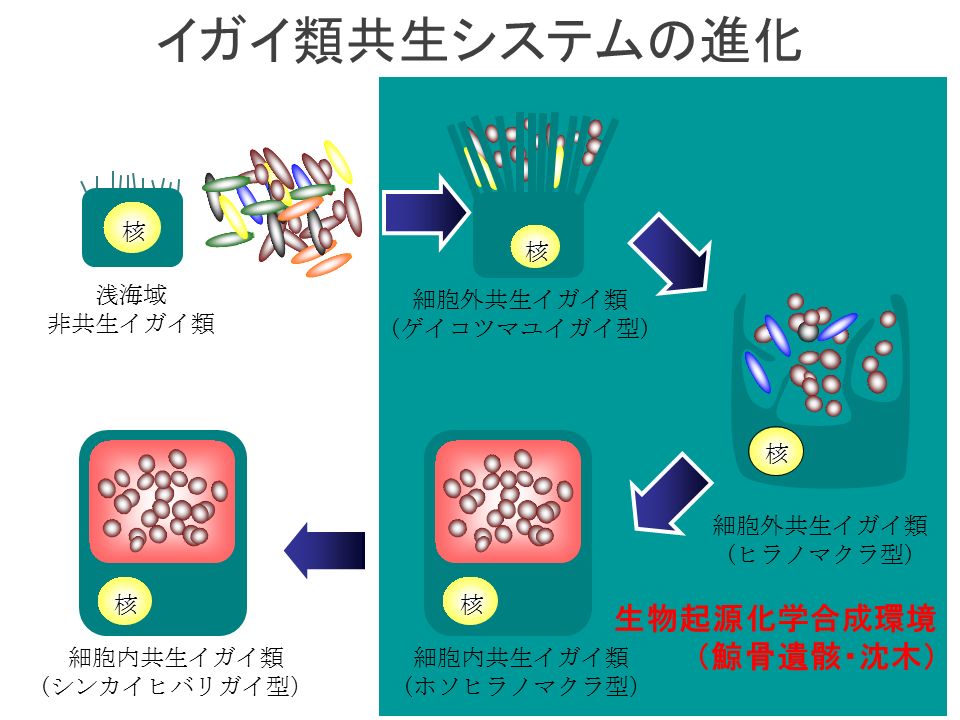
説明文の23
浅海域のイガイ類は化学合細菌と共生していない。ゲイコツマユイガイは細胞外で微絨毛に細菌を付着させ、ヒラノマクラは細胞膜が突き出して細胞外で細菌を囲みこんでいる。ホソヒラノマクラはシンカイヒバリガイ類と同様に細胞内に細菌を共生させている。このように化学合成細菌との共生の遷移が鯨遺骸で起こっていることも、”進化的飛び石仮説”を支持している。
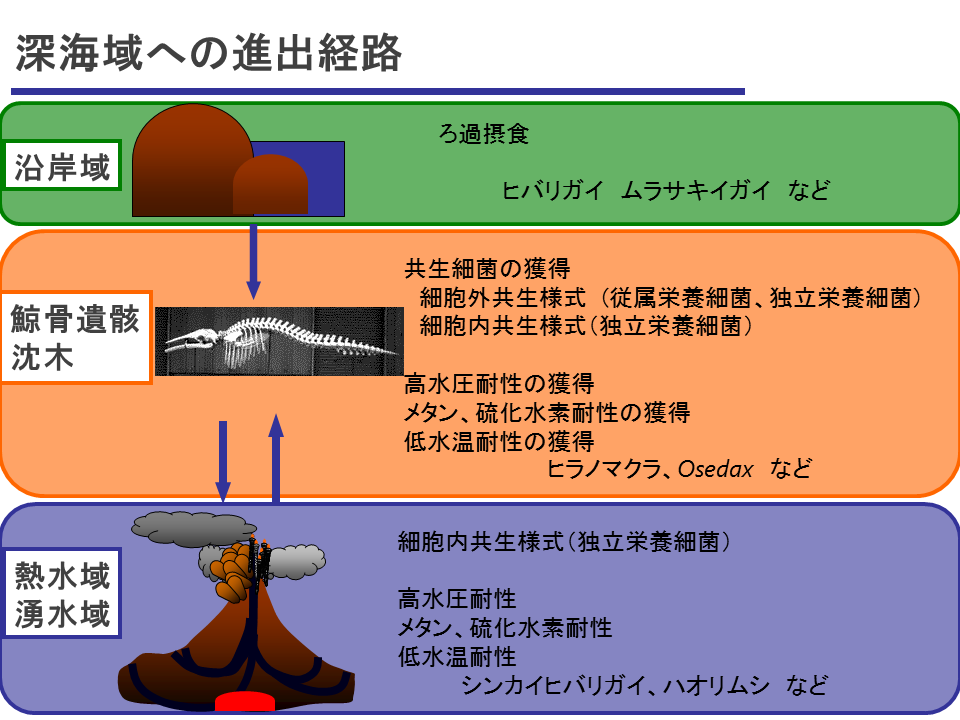
説明文の24
浅海の沿岸域のイガイ類のあるものが、様々な深度に散在する鯨遺骸や沈木に行きついた、そこで、高水圧に対する耐性や水温に対する耐性を徐々に身に付けていった。また、200m以深では光合成によるエネルギーが少なくなるので、光合成に頼らないエネルギー摂取の方法を身に付けなければならなかった。鯨遺骸や沈木はメタンや硫化水素を発生するので、それに対する耐性も身に付けることができた上、化学合成細菌とそこで出会うことになる。化学合成細菌との共生を細胞外から細胞内に徐々に洗練させ、とうとう熱水域や湧水域に進出し、化学合成生物群集の中で主要な位置を獲得した。