説明文の1
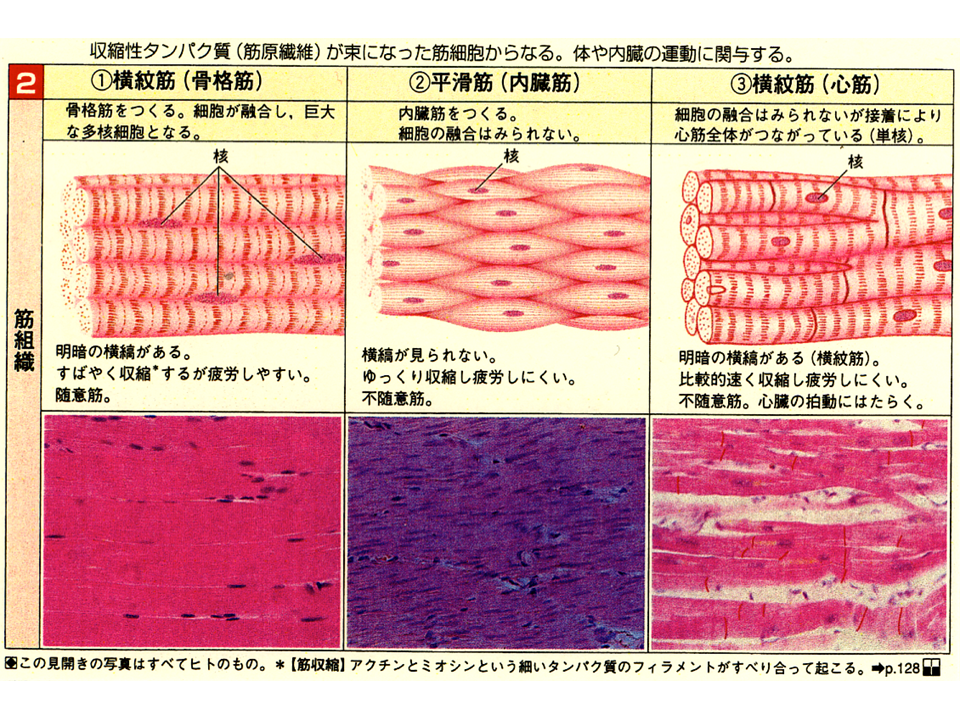
筋肉には大きく分けて3種類がある。骨格筋と内蔵筋、心筋である。筋肉を収縮させる蛋白質が緻密に秩序正しく配列しているため、骨格筋や心筋では縞模様がみえるため、横紋筋と呼ばれる。一方、内蔵筋は縞模様がみえないため、平滑筋とも呼ばれる。骨格筋は自分の意志で収縮させることができるので随意筋であり、心筋と内蔵筋は意志とは無関係に収縮調節されるので不随意筋である。骨格筋はいくつかの細胞が融合した巨大な多核(融合した細胞の核がたくさん入っている)細胞である。心筋や内蔵筋では細胞は融合することなく基本的に単核であるが、心筋ではいくつかの細胞が融合することがある。
説明文の2
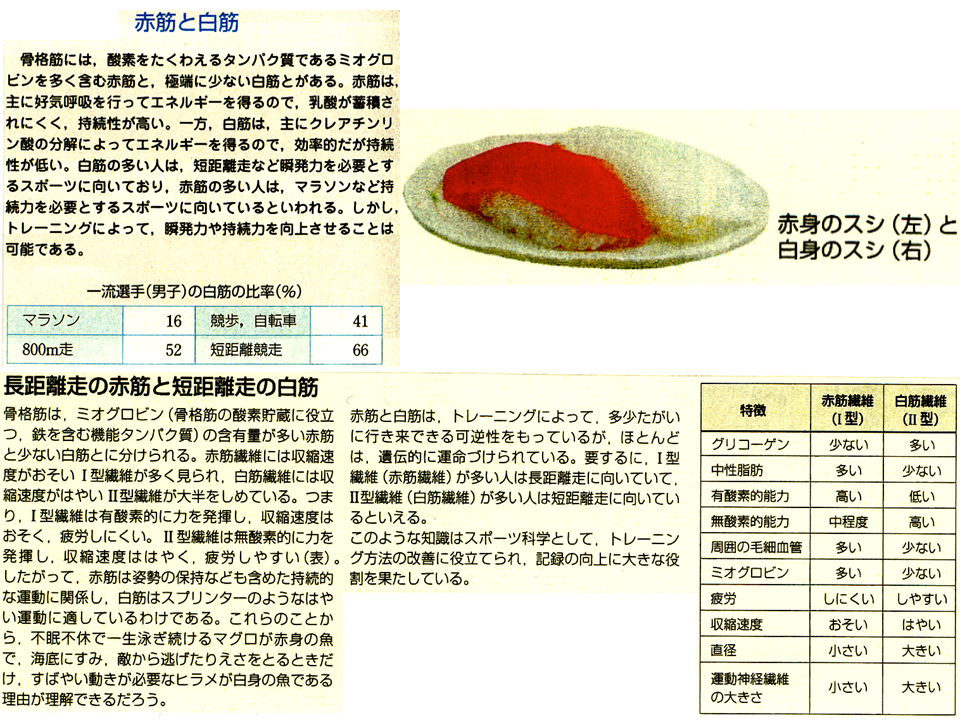
お寿司を食べる時、赤みの魚と白身の魚がある。色がなぜ違うのか?もちろん赤みの寿司ネタも、白みのものも筋肉である。赤みの筋肉はマグロのような常に泳いで筋肉を収縮させている魚に、白身の筋肉はヒラメのような普段はじっとしているのに、餌が近づくと瞬発的に筋肉を働かせる魚に発達している。すなわち色の違いは筋肉の性質の違いである。赤みの筋肉は遅筋とよばれ、持続的な運動に適している、白身の筋肉は瞬発的で力強い運動に適している。遅筋は、血液中のヘモグロビンに似た酸素と結合するミオグロビンという蛋白質を多く含んでいる。そのため血液と同じように赤っぽく見えるのである。同じ骨格筋なのにすいぶん違っているのがわかる。ただし、鮭の筋肉が赤色っぽいのは別の理由による。鮭の筋肉はもともと白である。また、敵が来ると瞬発的に翼を羽ばたいて逃げるニワトリのささみ(大胸筋)は白っぽい速筋であるが、いつも飛翔する鳥のささみは赤っぽい遅筋である。空を飛ぶためには持続的に筋肉を収縮させなければならない。

説明文の3
走るのがただ速ければ、マラソンと100m走のどちらでも金メダルを取ったアスリートがいるはずなのになぜいないのであろうか?2種目の練習をするのは時間的にも肉体的にも大変だから?それもあるだろう、がこれはもともと筋肉の質の違いによる。マラソンランナーは短い距離を100m走なみに走る必要はない、瞬発力は劣るが、疲れにくい持続的な運動に適した遅筋を持っている。一方、100mのスプリンターは長い距離を走る必要はない、疲れやすいが、瞬発力の必要な運動に適した速筋を持っている、どちらかに卓越しなければアスリートは勝つことができない。両方卓越した筋肉を持っている人がいたら、マラソンと100m走の両方勝てるのでは?それはない。マラソンをしている時速筋はいらない、ただ重荷になるだけである。逆に100m走をしている時遅筋はいらない、ただ重荷になるだけである。
説明文の4
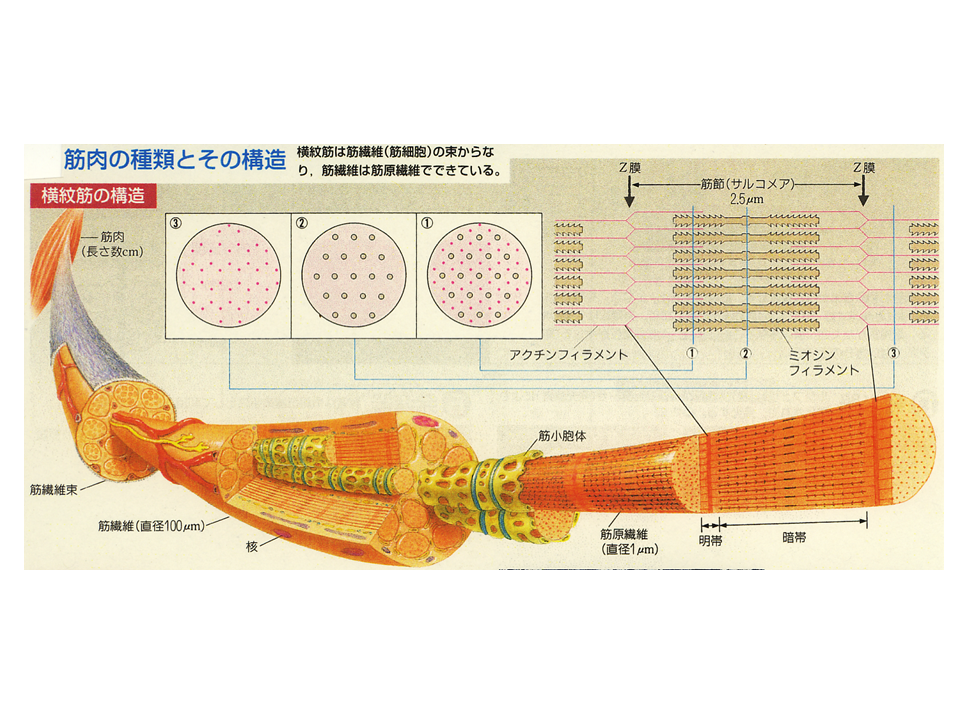
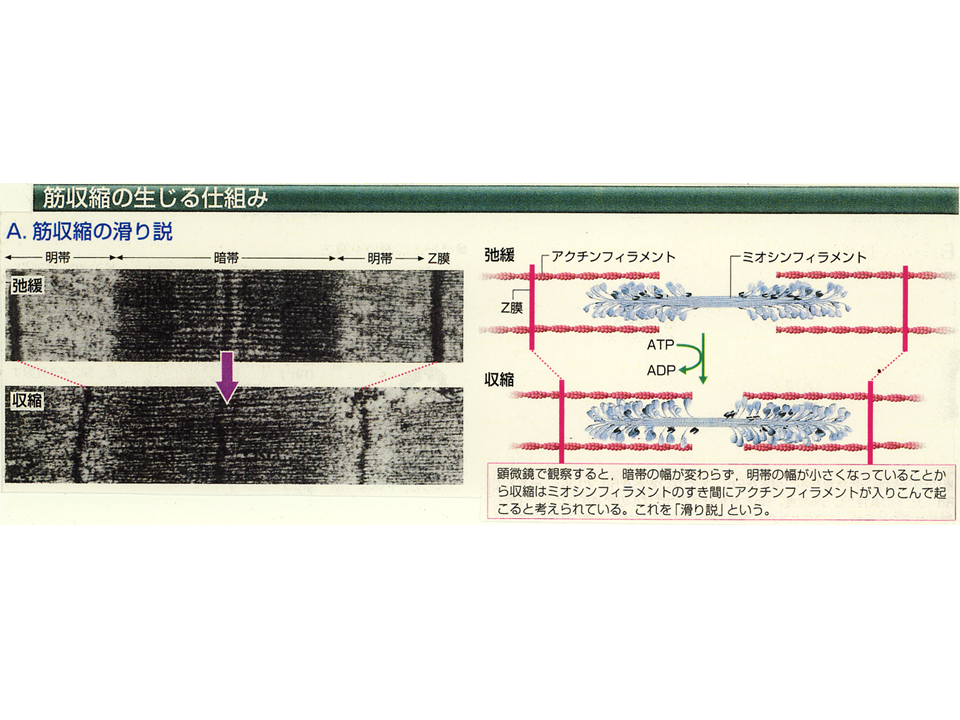
骨格筋を分解してみると、筋繊維が集まって束になっている(筋繊維束)。筋繊維は筋細胞のことであり、多数の細胞が融合して巨大な(細長い)細胞となっている。この筋繊維は筋肉を収縮させる蛋白質が規則的に並んだ筋原繊維を含んでいる。筋原繊維の基本的な単位は筋節(サルコメア)であり、Z膜が作るしきりの中に、主にアクチンから成る細い繊維(アクチンフィラメント)と主にミオシンから成る太い繊維(鬼の金棒のようにみえるミオシンフィラメント)が並んでいる。
説明文の5
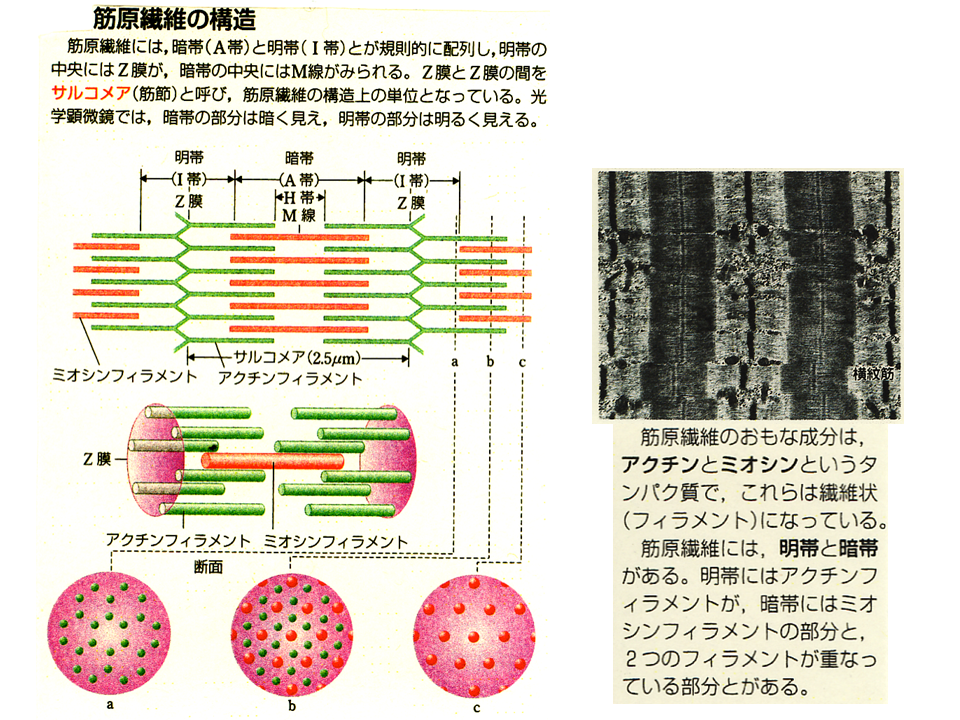
主にアクチンフィラメントがある所は明るく見えるので明帯、主にミオシンフィラメントがある所は暗く見えるので暗帯と呼ばれる。暗帯ではアクチンフィラメントとミオシンフィラメントが一緒に存在する所もある。
説明文の6
筋肉が収縮する時アクチンフィラメントがミオシンフィラメントの内側に引きずり込まれて(鬼の金棒のトゲトゲの部分がアクチンと結合して引っ張り込む)、サルコメアの長さが短くなる。そうすると筋原繊維の長さも短くなり、筋細胞の長さも短くなり、筋繊維束の長さも短くなるので、筋肉が収縮する。筋肉が収縮していない(弛緩している)時には、明帯がはっきりみられるが、収縮するとアクチンフィラメントが引きずり込まれるので、明帯があまり見えなくなる。
説明文の7
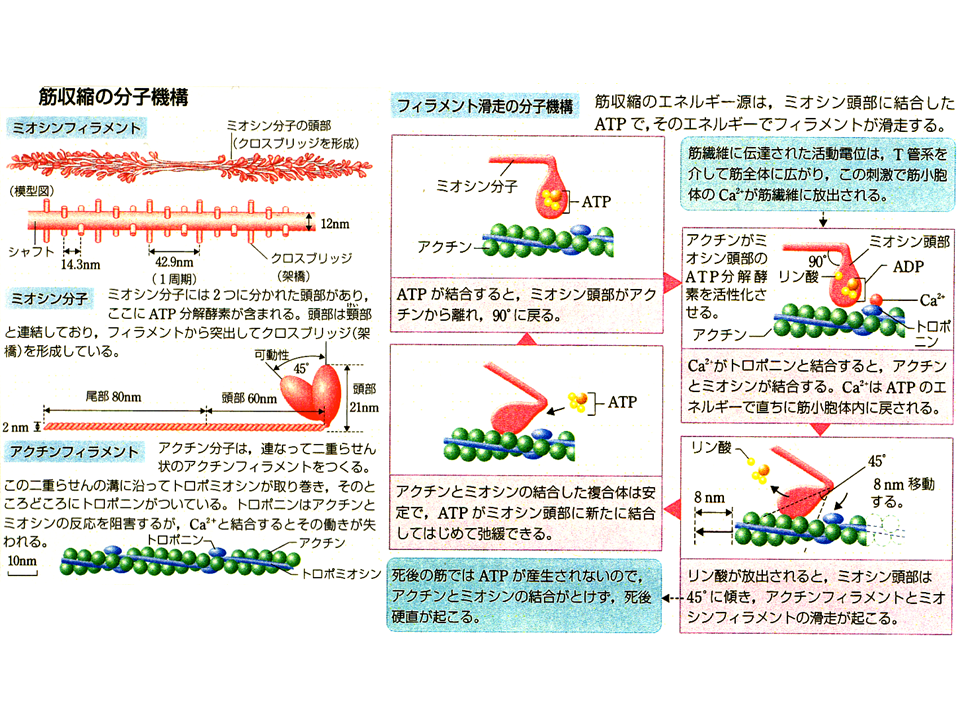
収縮はエネルギーの基であるATPを使いながら起こる。ミオシンはATPをADPとリン酸に分解しながらアクチンに結合し、ミオシン頭部の角度が変わることによってアクチンを引き込む。ミオシン頭部というのは、ミオシン分子の端っこで、鬼の金棒のトゲトゲにあたる部分である。筋肉の収縮・弛緩を調節するために必要なのは、アクチンとミオシンとATPだけではなく、カルシウムやトロポミオシンおよびトロポニンといった蛋白質も重要な役割をする。アクチンフィラメントがミオシンフィラメントにすべるように引くこまれる収縮過程を滑り説と呼ぶ。
説明文の8
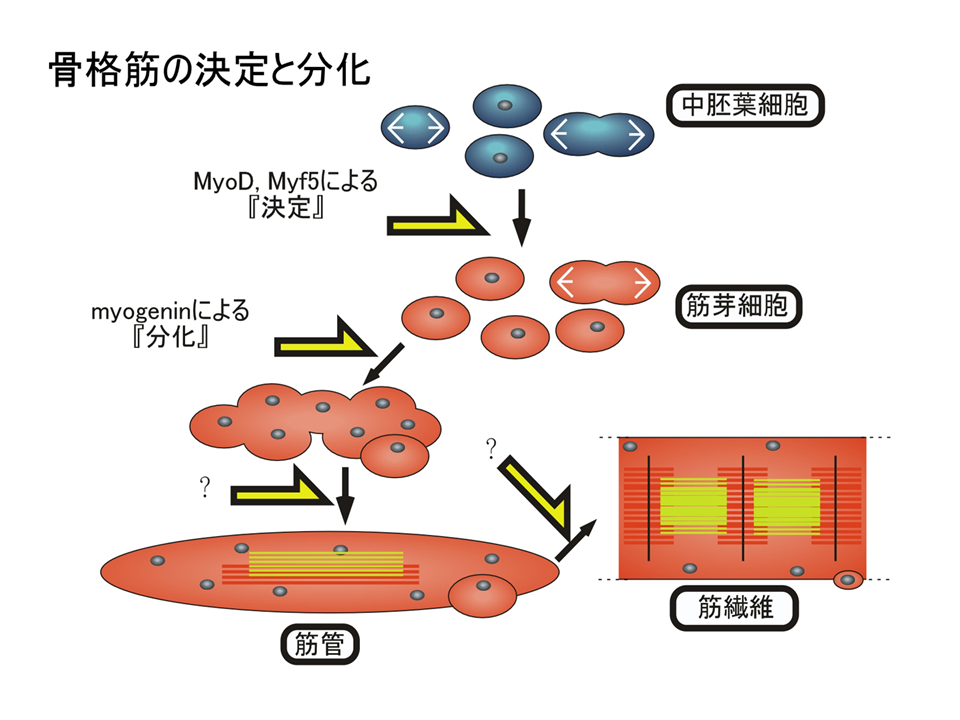
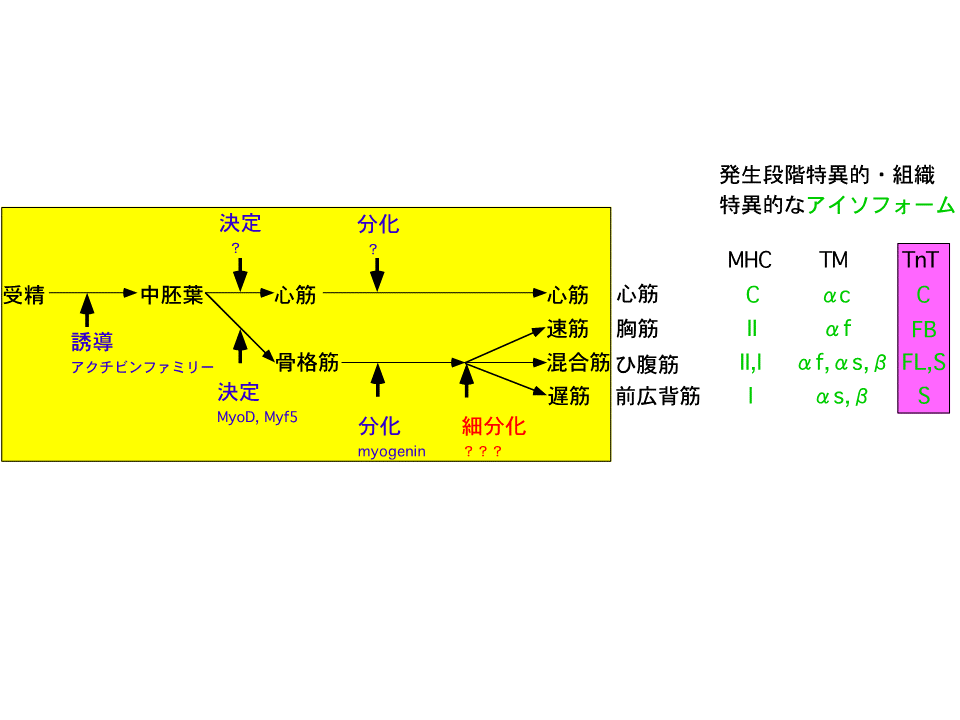
骨筋肉は昔から発生の材料として利用されてきた。それは、筋形成の過程で細胞融合や筋肉収縮装置の蛋白質が蓄積するので、非常にわかりやすいからである。このように細胞融合するような形成過程をたどるのはほぼ筋肉だけである。そのため、どのように筋肉ができるのかその機構も一番最初に明らかにされた。まず、受精卵が分裂していくと、大きく内胚葉、中胚葉、外胚葉という細胞のグループに分かれて行く。これらのグループは将来何になるか、その運命が違っている。中胚葉の細胞は筋肉になったり、骨になったりすることができる。中胚葉細胞の中で、MyoDあるいはMyf5という遺伝子が働くと、その細胞は筋肉に成るべく運命が決まる(筋芽細胞になる)。運命が決まるこの過程を「決定」という。さらにmyogeninという遺伝子が働くと、運命に従って実際に筋肉になっていく(筋管さらに筋繊維)。この過程を「分化」という。中胚葉細胞や筋芽細胞は分裂して増え、見た目で区別できない。筋管になる際、筋芽細胞は分裂をやめ融合する。
説明文の9
中胚葉細胞は中胚葉誘導によって形成される。どのように中胚葉誘導が起こるのか、その機構はかなり長い間わからなかった。しかし、日本人研究者(浅島先生)が世界で初めてそのヒントをつかんだ。アクチビンファミリー(今はTGF-β)という遺伝子のグループが働くと中胚葉細胞ができるのだ。そうすると筋肉は、中胚葉細胞の形成に始まり、最終的に筋肉ができるまですべてが明らかになったようにみえる、本当にそうだろうか?骨格筋がなぜ速筋や遅筋になるのか解明されていないではないか。速筋や遅筋の性質はミオグロビンの量だけでなく、筋肉の収縮・弛緩に関係する蛋白質の違いによって決まっている。その違いを生み出す機構は何であろうか。ヒトではほとんどの筋肉が速筋や遅筋の両方から成る混合筋である。しかし、ニワトリの大胸筋はほぼ100%速筋,背中の前広背筋はほぼ100%遅筋であり、速筋と遅筋の形成機構を調べるにはニワトリは良い材料である。
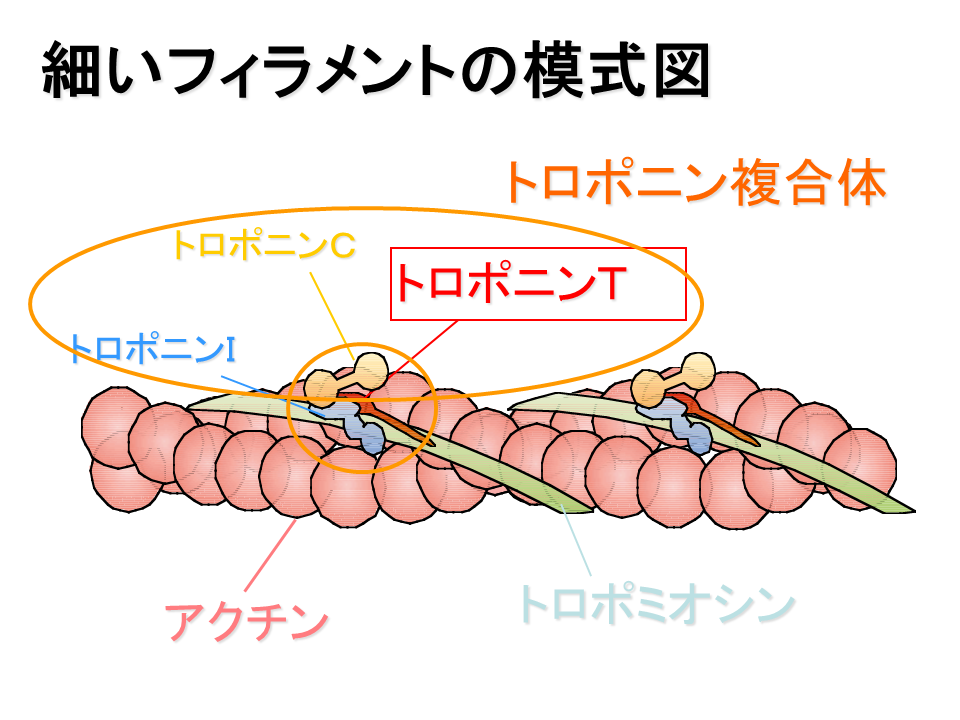
説明文の10
筋肉の収縮・弛緩にはトロポミオシンおよびトロポニンといった蛋白質が必要である。トロポミオシンは、球状のアクチンが繋がっているアクチンフィラメントにリボンのように巻き付いている。トロポニンは、そのトロポミオシンにのっかっている。トロポニンは3成分から成る。トロポニンTはトロポニンをトロポミオシンに結合させる役目をしている(トロポニンTのTはトロポミオシンtropomyosinのT)、トロポニンCはカルシウムと結合する(トロポニンCのCはカルシウムcalciumのC)、トロポニンIはカルシウムの細胞内の濃度が低い時にアクチンフィラメントとミオシンフィラメントの結合を阻害している(トロポニンIのIは阻害inhibitionのI)。トロポニンとその作用機構を明らかにしたのも日本人(江橋先生)である。骨格筋が意志(神経刺激)によって収縮する、その機構を分子レベルで明らかにした偉大な研究である。
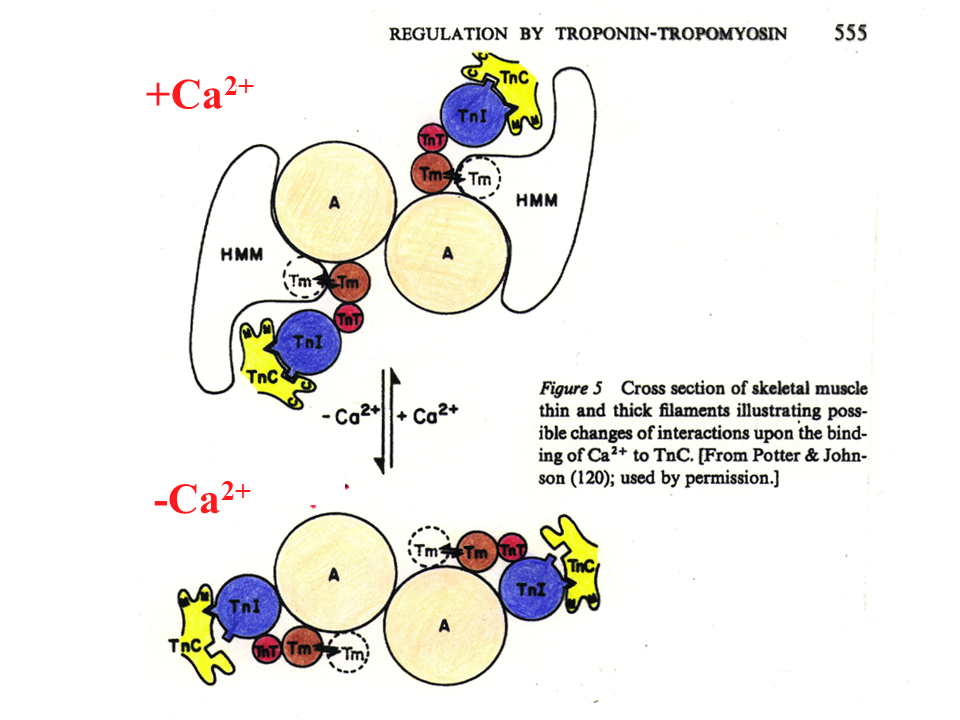
説明文の11
アクチンフィラメントの横断面をみてみると、神経刺激によって筋小胞からカルシウムが放出され、細胞内のカルシウムの濃度が高くなると(+Ca2+)、アクチンとミオシン頭部の(HMM)相互作用ができるようになり、筋は収縮する。神経刺激がなくなるとカルシウムは筋小胞に戻り、細胞内のカルシウム濃度が下がる(-Ca2+)。そうするとトロポニンIとトロポミオシンがアクチンとミオシン頭部の間に入り込んで相互作用を阻む。それで筋肉は弛緩する。トロポニンIとトロポミオシンがアクチンとミオシン頭部の相互作用を阻害しているので、これを立体阻害モデルという。
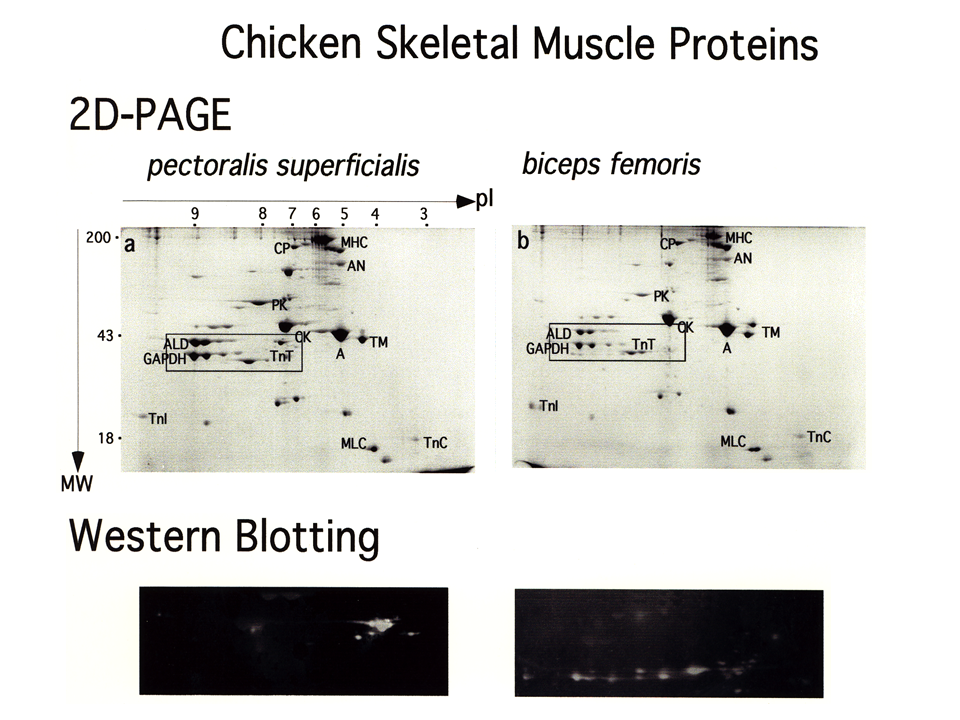
説明文の12
筋肉収縮に重要なトロポニンに注目する。蛋白質を包括的に調べることができる2次元電気泳動(2D-PAGE)でニワトリの速筋である大胸筋(pectoralis superficialis)と大腿二頭筋(biceps femoris)を調べると、トロポニンT(TnT)に何となく違いがみられれる(四角の枠で囲った部分)。速筋型トロポニンTに対する抗体を用いて、トロポニンTだけをクローズアップしてみると(Western Blotting)、トロポニンTはいくつもの成分を含んでおり、しかも2つに筋肉の間で明らかにその成分が異なる。この成分をアイソフォームという(基本的に同じ機能を果たすが構造や性質が違う蛋白質)。この2次元電気泳動パターン上には、非常に分子量の大きいミオシン(MHC)も、分子量が小さく非常に酸性の(等電点の小さい)トロポニンC(TnC)も、非常にアルカリ性の(等電点の大きい)トロポニンI(TnI)も、アクチン(A)やトロポミオシン(TM)もみえる。筋肉収縮・弛緩にかかわる多くの蛋白質を網羅的に調べることができる2元電気泳動法は。私の師匠の平林民雄が開発したものである。
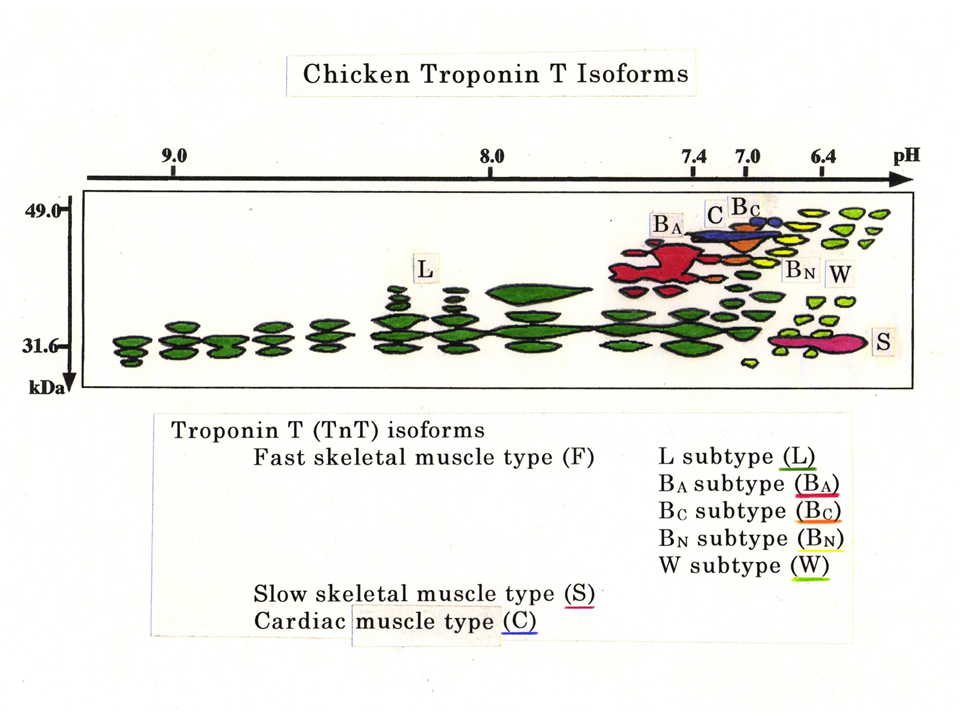
説明文の13
2次元電気泳動法と抗体を使ったWestern Blottingによって、ニワトリのトロポニンTにはこれほど多くのアイソフォームがあることがわかった。緑色は主に下肢の筋肉でみられる速筋型(L型)、赤色は大人の胸筋にみられる速筋型(BA型)、橙色は主にヒヨコの胸筋にみられる速筋型(BC型)、黄色は主に孵化したばかりの時にみられる速筋型(BN型)、黄緑色は上肢(翼)だけにみられる速筋型(W型)、赤紫色は遅筋型(S型)、青色は心筋型(˙C型)。こんなにたくさんのトロポニンTアイソフォームがあることはなかなか信じてもらえなかった。ただいくつかのアイソフォームが徐々に分解されているだけはないかとみなされた。
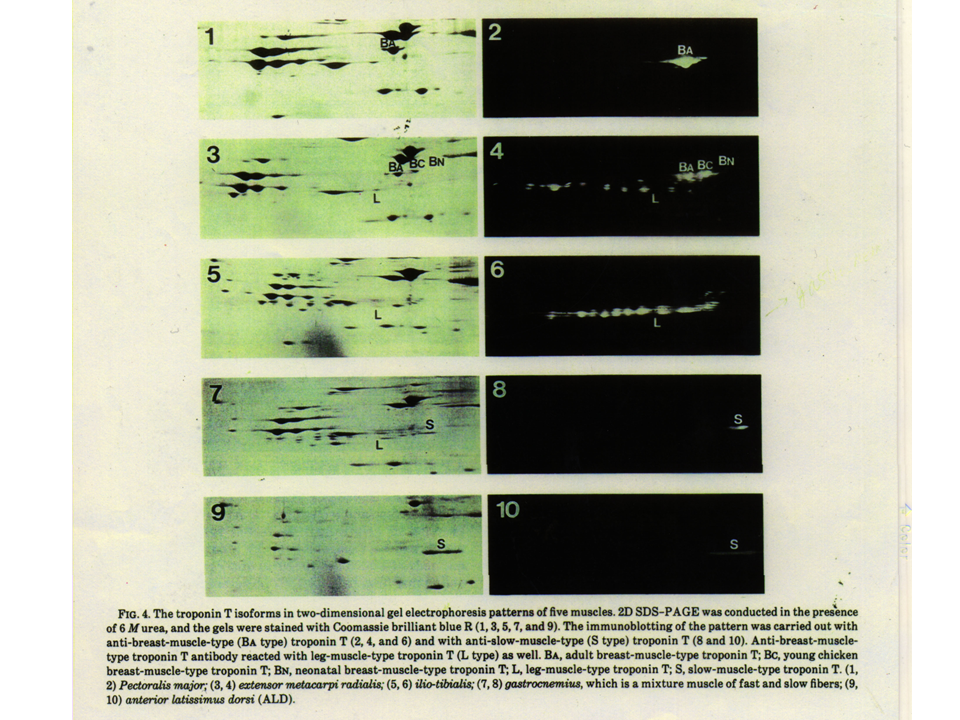
説明文の14
ニワトリの大人の筋肉のトロポニンTアイソフォーム。速筋の胸筋ではBA型(1と2)、速筋の上肢の筋肉ではL、BA、BC、BN(3と4)、混合筋の下肢の筋肉ではL型(5と6)とS型(7と8)、遅筋の前広背筋ではS型(9と10)、がみられる。なぜこのように筋肉によってトロポニンTアイソフォームが異なるのであろう?トロポニンTは筋肉の収縮・弛緩に重要な蛋白質である。これが異なるということは筋肉の収縮特性も異なるということであろうか?
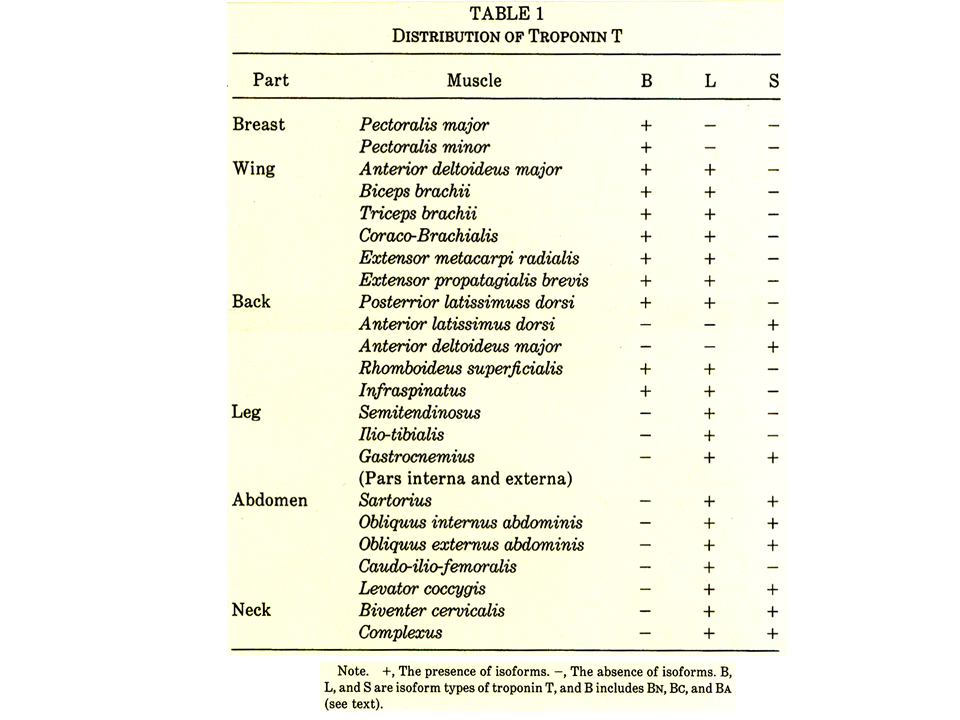
説明文の15
大人の胸、上肢、背中、下肢、腹、首の筋肉でどのようなトロポニンTアイソフォームがみられるかまとめたものである。Bは速筋型BA、BC、BNのすべてあるいはいずれかかがみられることを示している。Lは速筋型。Sは遅筋型。
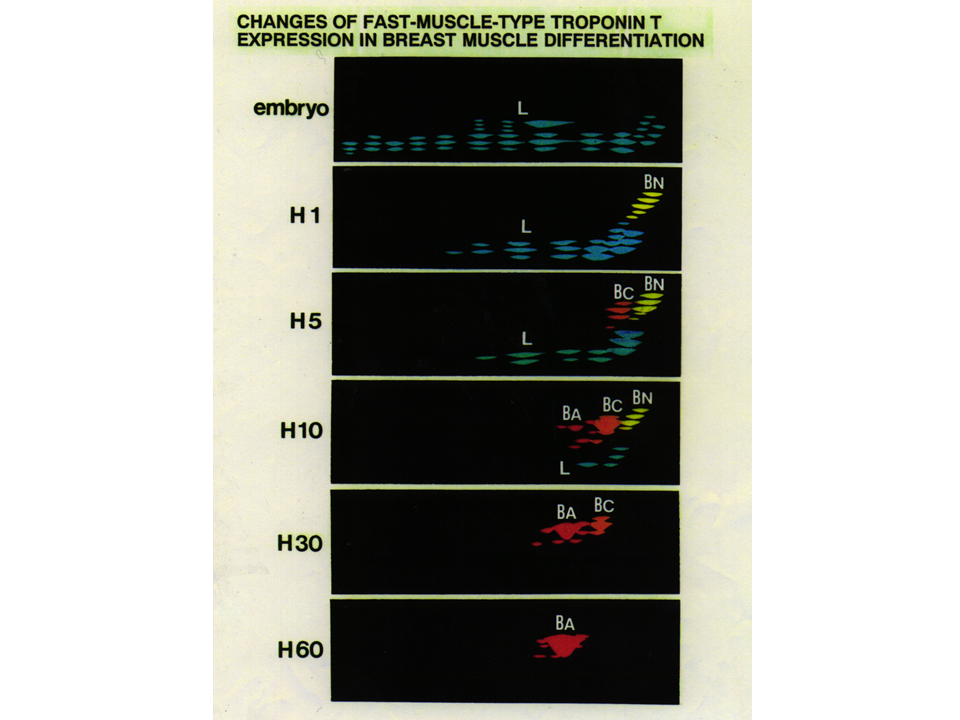
説明文の16
トロポニンTアイソフォームは発生に伴って変化する。大胸筋では、孵化する前はL型、孵化前後にBN型がみられるようになる。次にBC型がみられるようになり、最終的にBA型が現れて、大人ではBA型だけとなる。その間にL型、BN型、BC型は徐々に消えていく。L型はもともと大人の下肢でみつかったので、legのL、B型はもともと胸肉でみつかったので、brestのB。BN型のNは新生児neonatal、BC型のCはchick、BA型のAはadultを意味する。
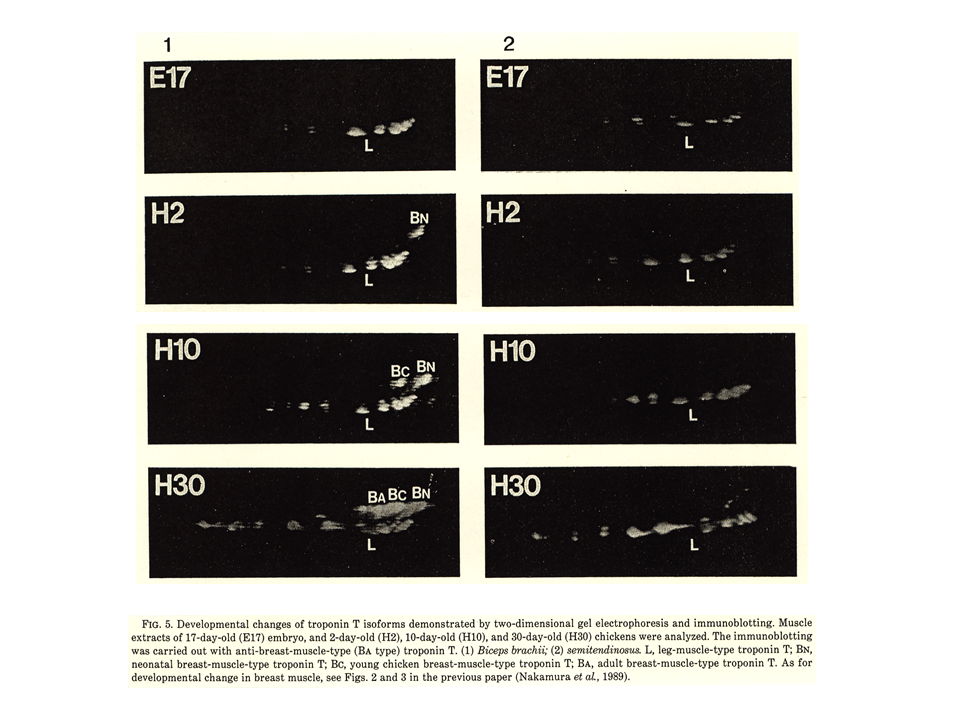
説明文の17
左の上肢の筋肉では、胸筋と同じように発生が進むが、L型、BN型、BC型は消えることなく、全部が大人でも残ってしまう。下肢の筋肉では、大きな変化はなくB型は現れない。E17は卵が産まれてから17日目、ニワトリは一般に21日目に孵化する。Eは胚embryo、Hは孵化hatch-out後を示す。発生に伴ってトロポニンTアイソフォームが変化するのはなぜか?各々の発生段階で筋肉に異なる収縮特性を与えるためではないか。
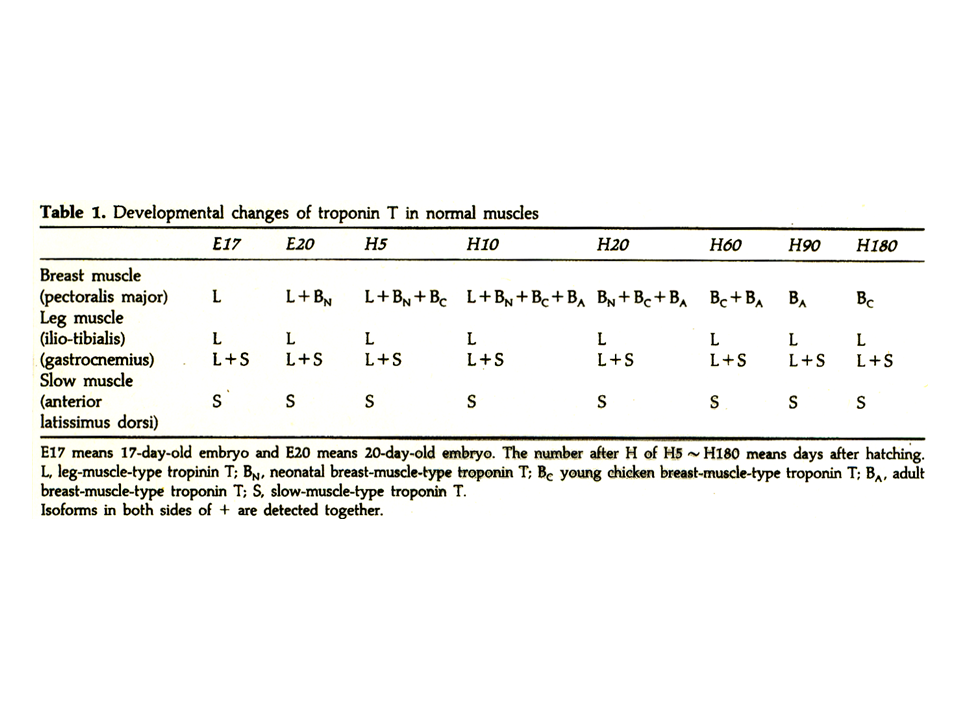
説明文の18
発生に伴ってトロポニンTアイソフォームがどのように変化するかをまとめたものである。胸の速筋では、孵化する前はL型、孵化前後にBN型がみられるようになる。次にBC型がみられるようになり、最終的にBA型現れて、大人ではBA型だけとなる。下肢の混合金では、ずっとL型とS型がみられる。背中の遅筋では、ずっとS型がみられる。
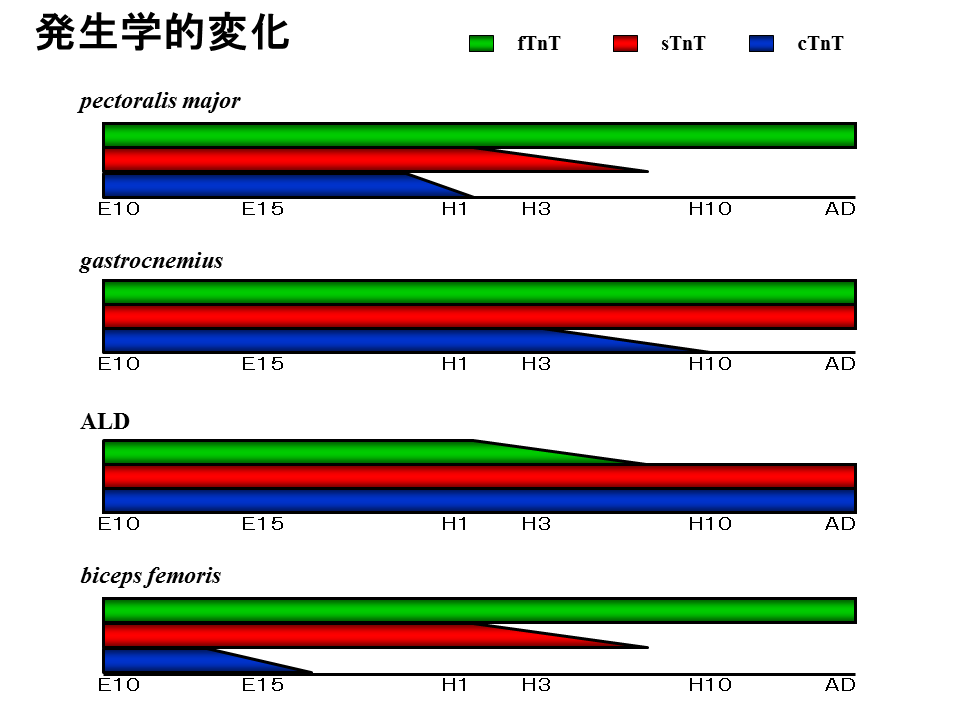
説明文19
もっと詳しく調べてみると、胸の速筋(pectoralis major)や下肢の速筋(biceps femoris)では、孵化前後まで遅筋型(sTnT)だけでなく、心筋型(cTnT)さえもみられる。下肢の混合筋(gastrocnemius)でも、孵化前後まで心筋型がみられる。さらに背中の遅筋(ALD)では、孵化前後まで心筋型だけでなく、速筋型(fTnT)さえもみられる。孵化前後までトロポニンTアイソフォームの遺伝子はきちんと調節されていないのかもしれない。
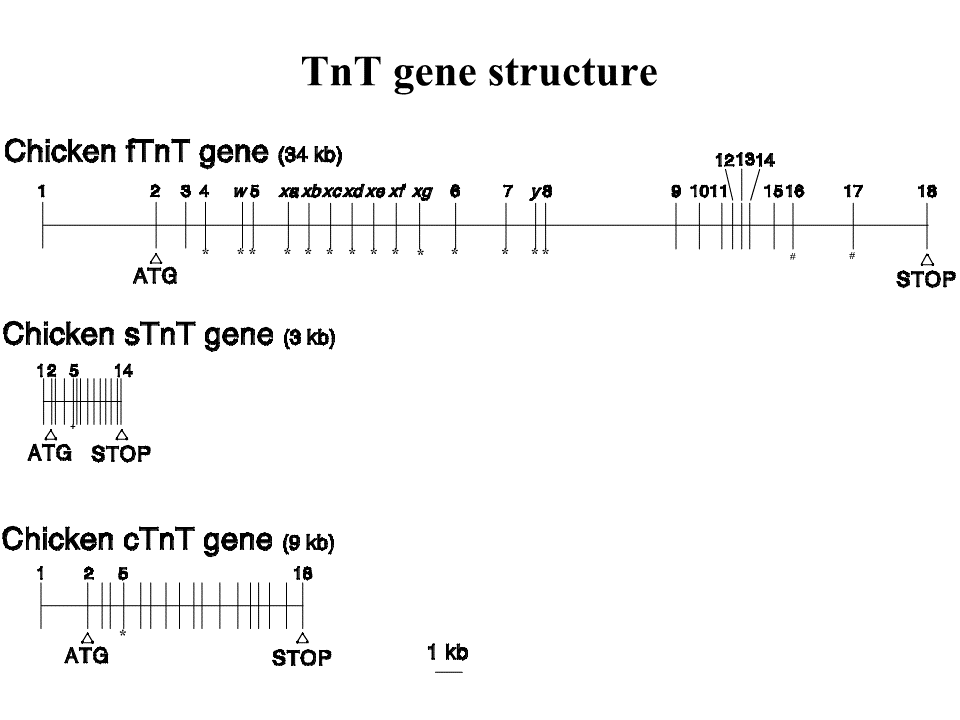
説明文の20
なぜこんなにたくさんのトロポニンTアイソフォームがあるのだろう。多くの脊椎動物には3つの遺伝子があることがわかっている。速筋(fast)型遺伝子、遅型(slow)型遺伝子、心筋(cardiac)型遺伝子である。速筋型遺伝子は34kbの長さがあり、27個のエクソン(楯の線で示されている)がある。遅型遺伝子は3kbの長さしかなく、エクソンも14個しかない。心筋型遺伝子は10kbの長さがあり、18個エクソンがある。これら3つの遺伝子はもともと1つ祖先遺伝子から進化したとされる。本当なのだろうか?なにせ遅型遺伝子は速筋型遺伝子のエクソン1と2の間におさまってしまうくらいの長さしかないのだ。
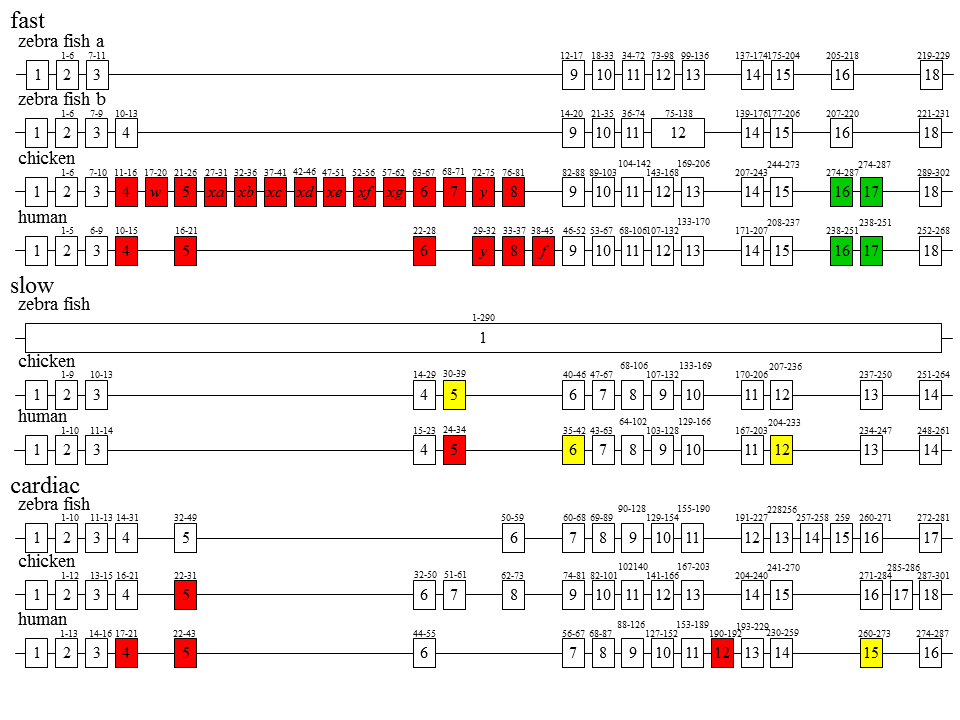
説明文の21
さらに驚くべきことがわかった。色で示した四角は選択的スプライシングを受けるエクソンである。硬骨魚類であるzebra fishでは、速筋型遺伝子が2つあるらしい。また、選択的スプライシングが起こらない上、遅型遺伝子はエクソン・イントロン構造がないらしい?ヒトの遺伝子はニワトリと同じく3つであるが、速筋型遺伝子ではスプライシングされるエクソンの数がニワトリより少ない。赤色はカセット型スプライシングを受けるエクソンで、緑色は相互排他的スプライシングを受けるエクソン、黄色は選択的3‘スプライス部位をもつエクソンである。
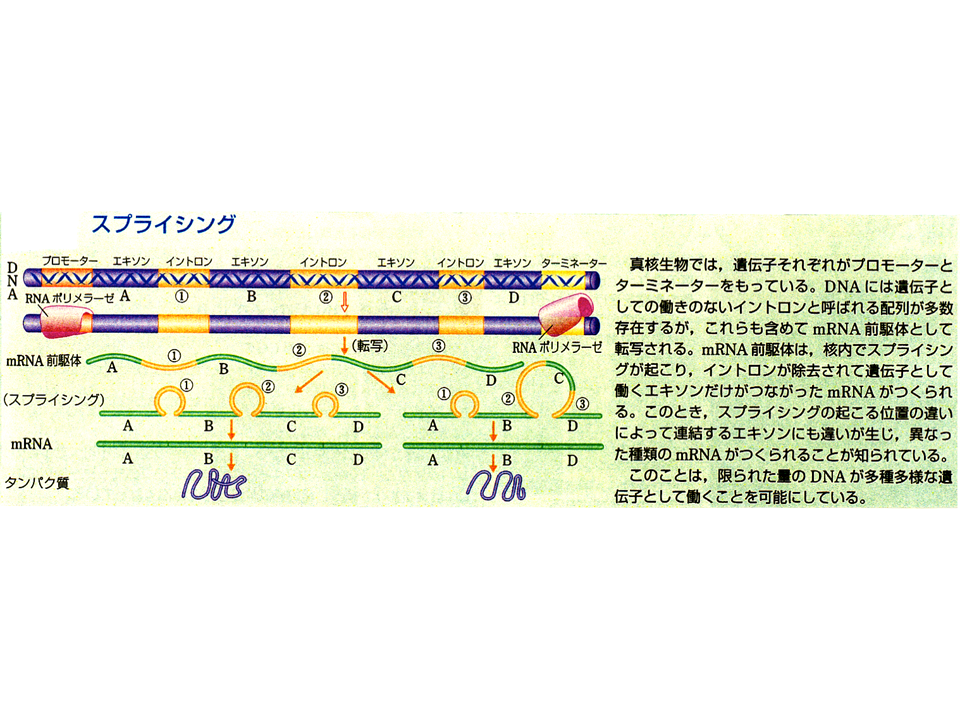
説明文の22
選択的スプライシングとは何であろうか。核のDNAの情報をそのままコピーしたのがmRNA前駆体である。この前駆体の中の蛋白質を作り上げるのに重要な部分であるエクソンをつなげ、いらない部分であるイントロンを切り取るのがスプライシングであり、これによってmRNA前駆体がmRNAとなる。そのスプライシングの際、いる部分といらない部分の選択のやり方を変えるのが選択的スプライシングである。例えば、A、B、C、Dの4つのエクソンをすべてを含むmRNAとA、B、Dの3つのエクソンしか含まないmRNAが作られる場合がある。前者ではエクソンCは必要な部分として、後者では不必要な部分(つまりイントロン)として選択されたことになる。
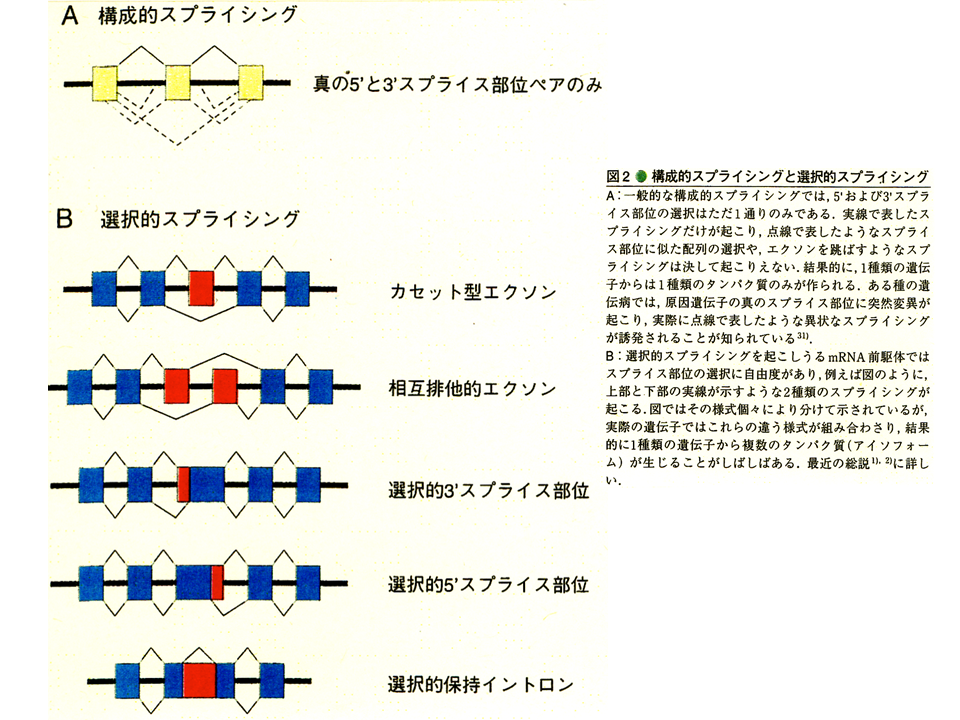
説明文の23
選択的なスプライシングにはいろいろなやり方がある。トロポニンT遺伝子でみられた、カセット型は赤色のエクソンを選択するか、しないかで2通りのmRNAができる可能性がある。相互排他的な場合には、赤色のエクソンのどちらかが必ず選択されるので、これも2通りのmRNAができる可能性がある。選択的3‘スプライス部位をもつ場合は、エクソンの前端の赤色の部分を取るか取らないかでやはり2通りのmRNAができる可能性がある。すなわち、カセット型と選択的3‘スプライス部位をもつ場合は1つのエクソン(削除される場合はイントロンとなるが)が関係して、2種類のmRNAが、相互排他的ば場合は2つのエクソンが関係して2種類のmRNAができることになる。
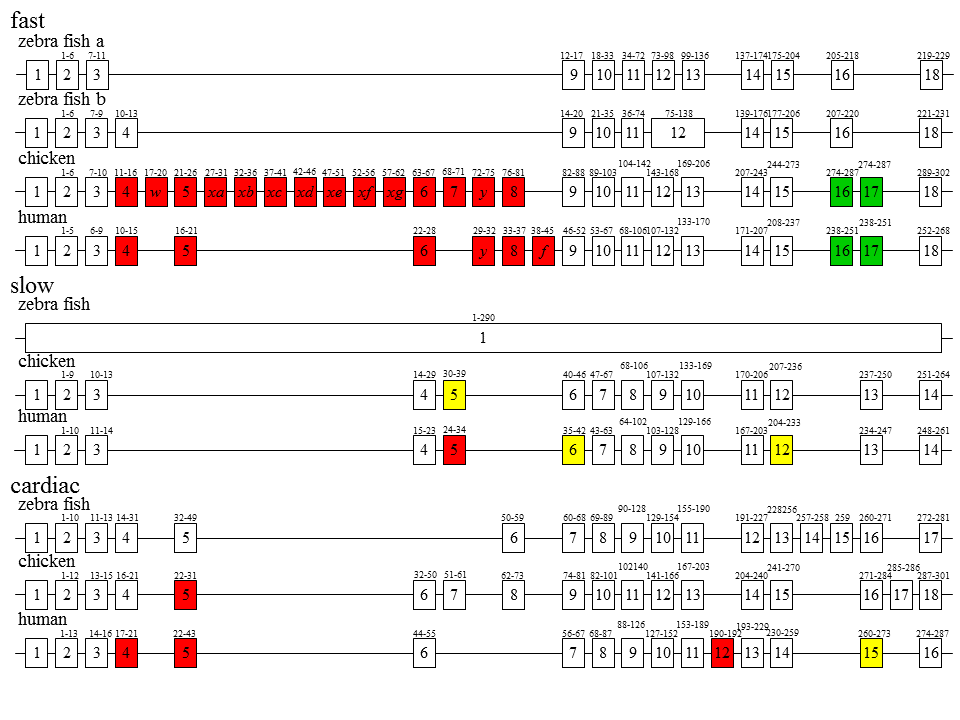
説明文の24
ニワトリの遺伝子をみてみると、遅筋型遺伝子は選択的3‘スプライス部位をもつエクソンが1つあるので、2種類のmRNA(すなわち2種類のトロポニンTアイソフォーム)が作られる可能性がある。 心筋型遺伝子はカセット型エクソンを1つもつので、2種類のmRNAが作られる可能性がある。問題は速筋型遺伝子である。カセット型エクソンが14個あり、1個で2種類のmRNAを作ることができるので、これだけで214種類のmRNAを作ることができる。さらに相互排他的エクソンが1つあるのでここでも2種類のmRNAを作ることができる。全部組み合わせると1つの遺伝子から最大で215(約3万)のトロポニンTアイソフォームを生み出すこができるのである。このようにな遺伝子は私が知る限り他にない。
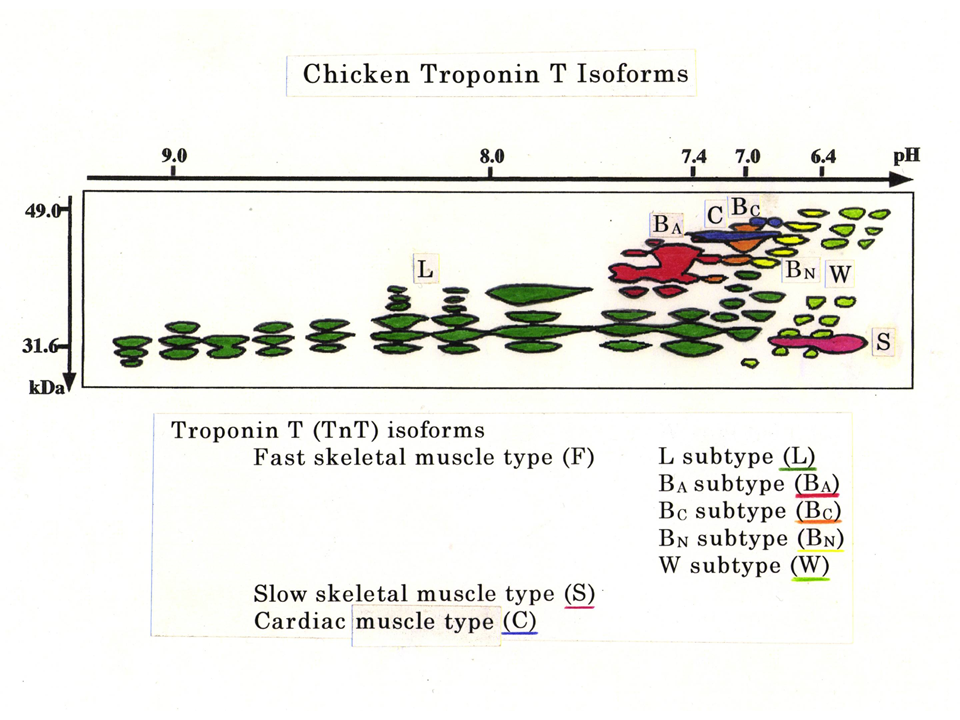
説明文の25
蛋白質レベルでたくさんのトロポニンTアイソフォームがみつかり、このほとんどは分解産物であろうと疑われた。しかし、蛋白質分解酵素を抑制して注意深く実験をおこなっても、同じだけトロポニンTアイソフォームがみられるのある。また、1つの遺伝子から最大で約3万種類のトロポニンTアイソフォームが生み出されることが今や明らかになったので、ここにみられるトロポニンTアイソフォームは分解産物ではなく、遺伝子の選択的スプライシングから十分に説明できるものである。