説明文の1
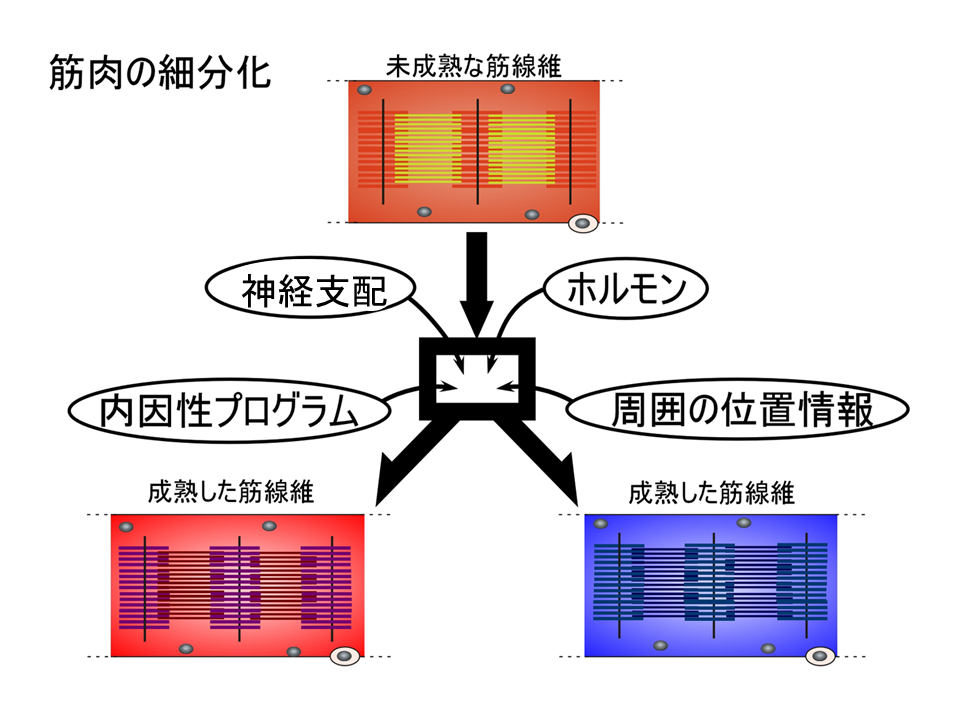
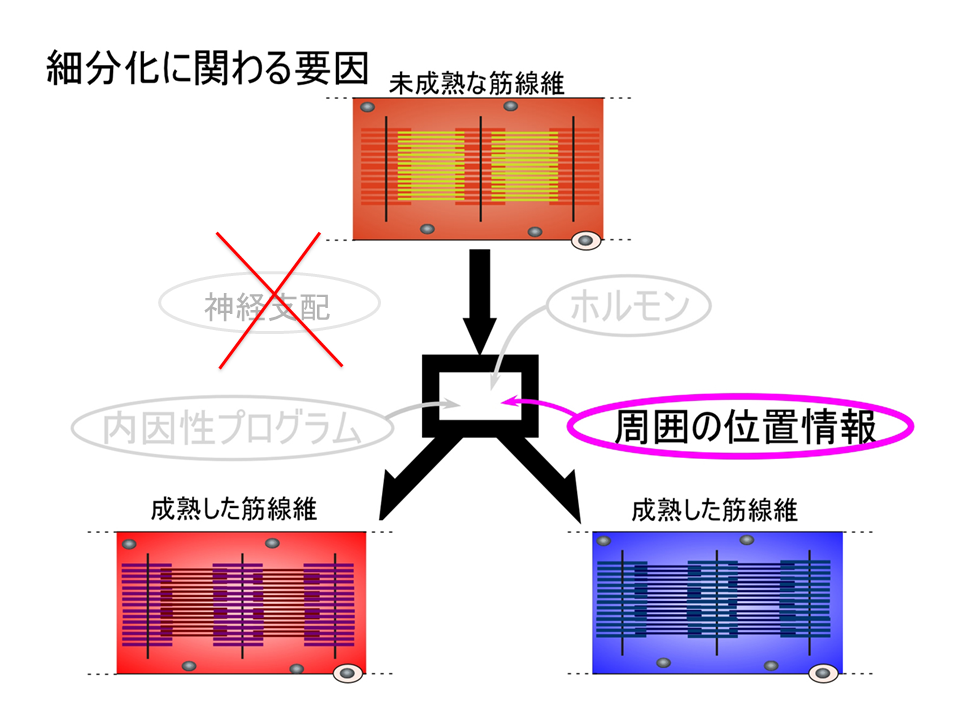
どのようにして速筋や遅筋、混合筋は形成される(すなわち細分化される)のだろうか?これらの骨格筋は、各々特徴的な収縮・弛緩を調節する蛋白質のアイソフォームをもつが、未成熟な筋肉ではアイソフォームの発現調節は厳密ではない。筋が成熟する過程で、特徴的なアイソフォームをもつよう調節する要因として、最初に1)ホルモン、2)神経支配、3)周囲の位置情報、4)内因性プログラムの、4つを考えた。

説明文の2
筋肉増強剤で知られるように、ホルモンは筋肉の形成に大きな影響を与える。しかし、ホルモンは血液で全身に運ばれるので、特定の部位で速筋や遅筋、混合筋に未成熟の筋肉を細分化させるのは難しいのではないか。もし、ホルモンが細分化に関わるとしても、ホルモンを受け取る筋肉側(ホルモンのリセプター)が既に違っていないと細分化は起こらない。つまり、ホルモンの影響を受ける前に既に筋肉側に違い(細分化)がなければならい。このように考えて、ホルモンの可能性をひとまず消し、神経支配、周囲の位置情報、内因性プログラムを考えることにした。
説明文の3
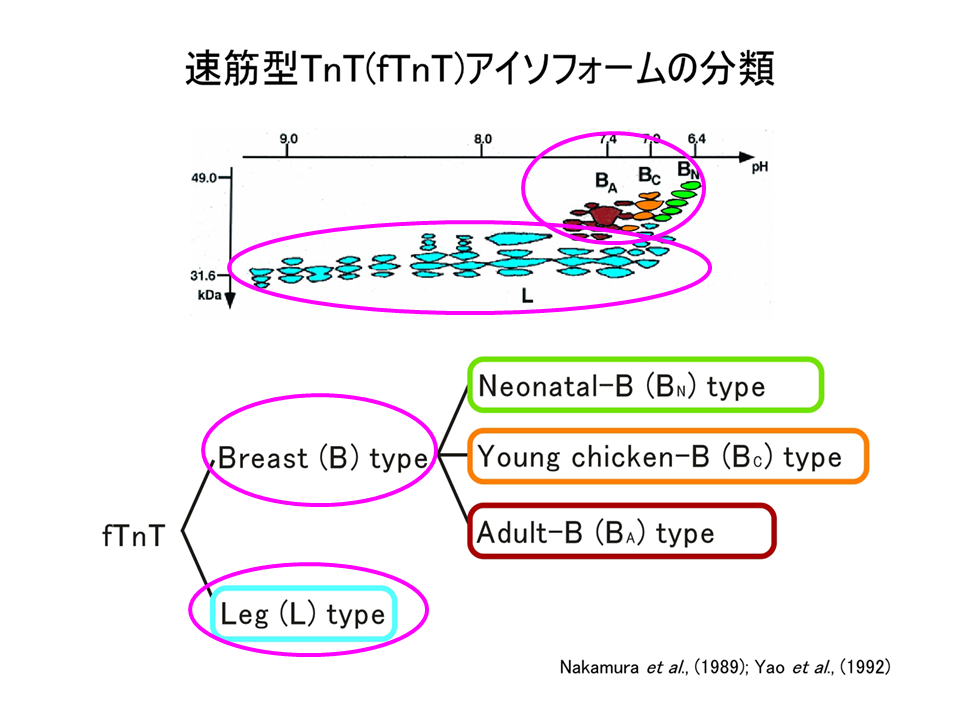
筋肉の細分化を探るために、トロポニンTを分子マーカーとして用いることにした。速筋型トロポニンT(fTnT)は、その遺伝子の中に多くの選択的エクソンをもっているため、1つの遺伝子から多数のアイソフォームが作られる。二次元電気泳動像パターンで示されるように、トロポニンTのB型とL型を比べると、B型は相対的に分子量が大きく、等電点はpH7付近にあるが L型は分子量が小さく、等電点は広い範囲にわたり、二次元電気泳動パターン上で明確に区別できる。
説明文の4
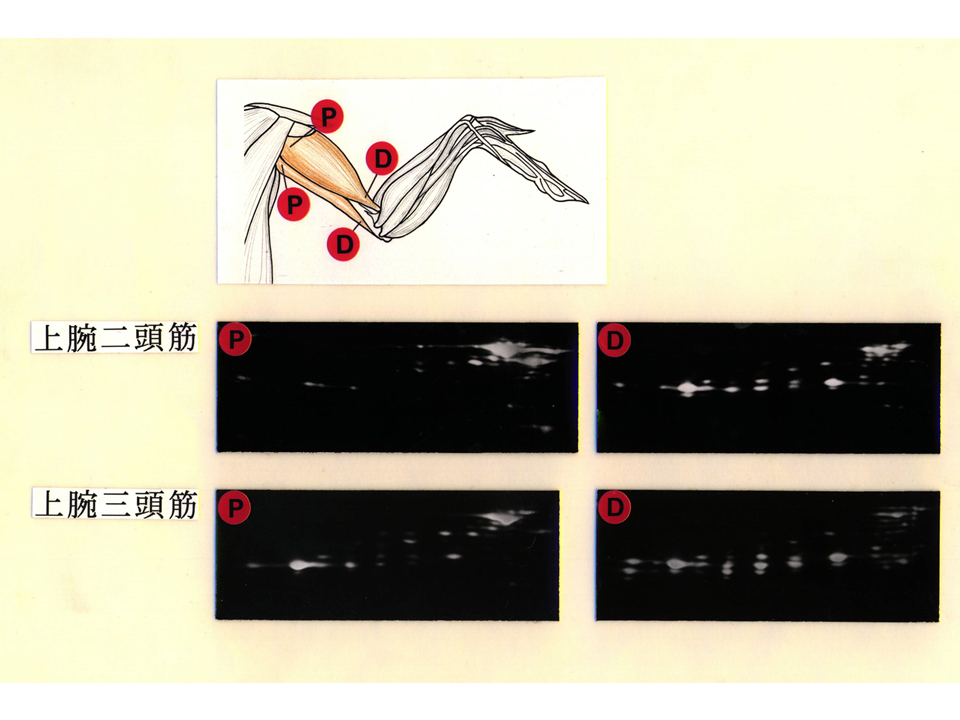
筋肉としては上肢の筋肉を用いることにした。上腕二頭筋と上腕三頭筋の速筋型トロポニンTアイソフォームのパターンを調べたところ奇妙なことがわかった。各々の筋肉の肩側ではB型の方がL型より多く、肘側ではL型の方がB型より量が多いように見える。各々の筋肉では、神経刺激に対して同調的に反応しなければならないが、収縮・弛緩を調節するトロポニンTアイソフォームに部位による違いがあっても良いのだろうか?発想を逆転させて、違いがあるからこそ同調的に働くことができるのだと考えることにした。
説明文の5
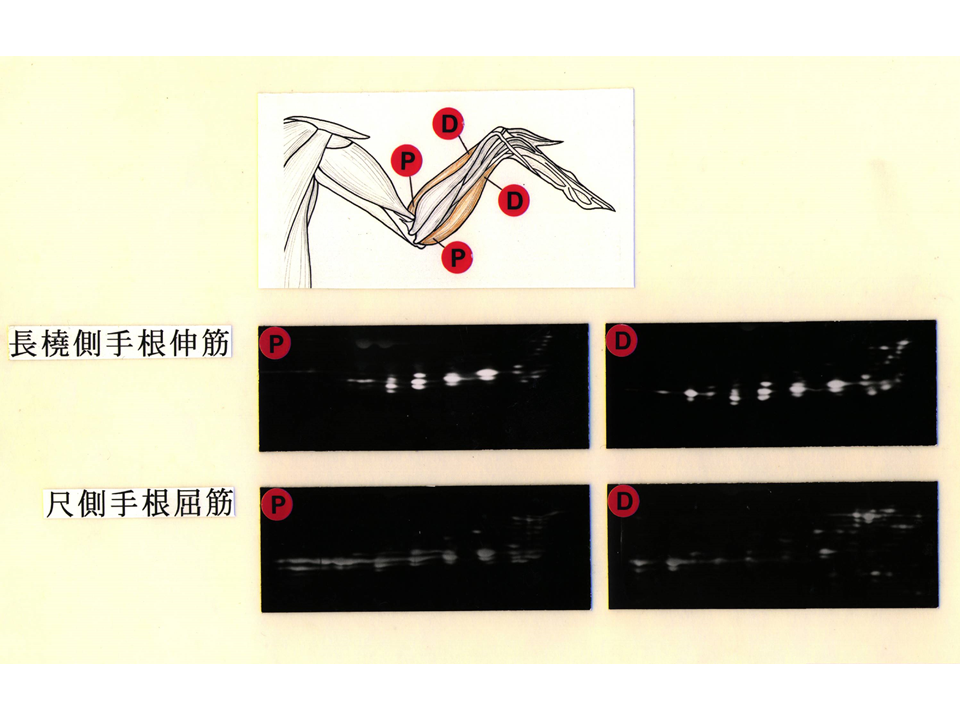
上腕に比べて、下腕の筋肉は複雑である。いくつかの筋肉を調べてみると、やはり肘側と手首側にトロポニンTアイソフォームの違いがあった。しかし、上腕の筋肉ほどではないようにみえる。このようにトロポニンTアイソフォームの発現に部位による違いがあるということは、速筋や遅筋、混合筋の細分化よりさらに細かい細分化があるのかもしれない。
説明文の6
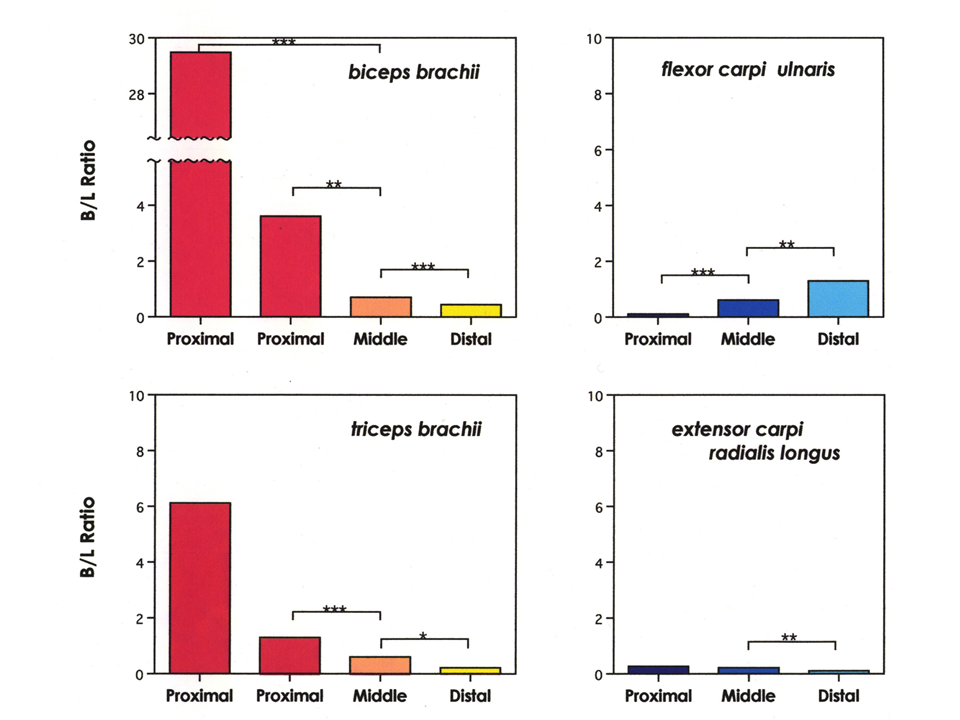
上腕二頭筋(biceps brachii)、上腕三頭筋(triceps brachii)、下腕の2つの筋肉で、両側と真ん中部分のトロポニンTアイソフォームのB型とL型の量を測定した。上腕二頭筋と上腕三頭筋では、B型に対するL型の量比(B/L)は、肩側(Proximal)、真ん中(Middle)、肘側(Distal)の順に小さくなっていく。肩側で2つの値があるのは大きな個体差がみられたためである。下腕の筋肉では、上腕と逆のパターンを示すもの、部位によって大きな違いを示さないものがあった。
説明文の7
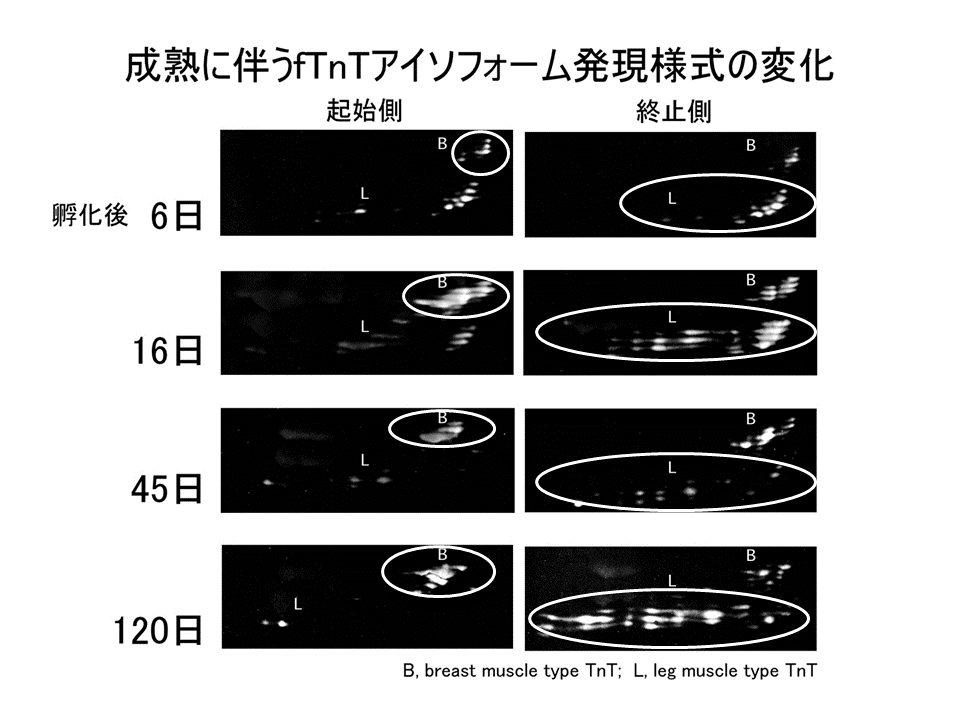
トロポニンTアイソフォームの発現の、1つの筋肉内の部位による違いは、発生に伴って現れてくる。孵化後6日目では、肩側(起始側)と肘側(終止側)の間に違いは見られなかった。孵化後16日目、45日目、そして120日目と、筋が成熟するに従って、違いがはっきりみられるようになった。
説明文の8
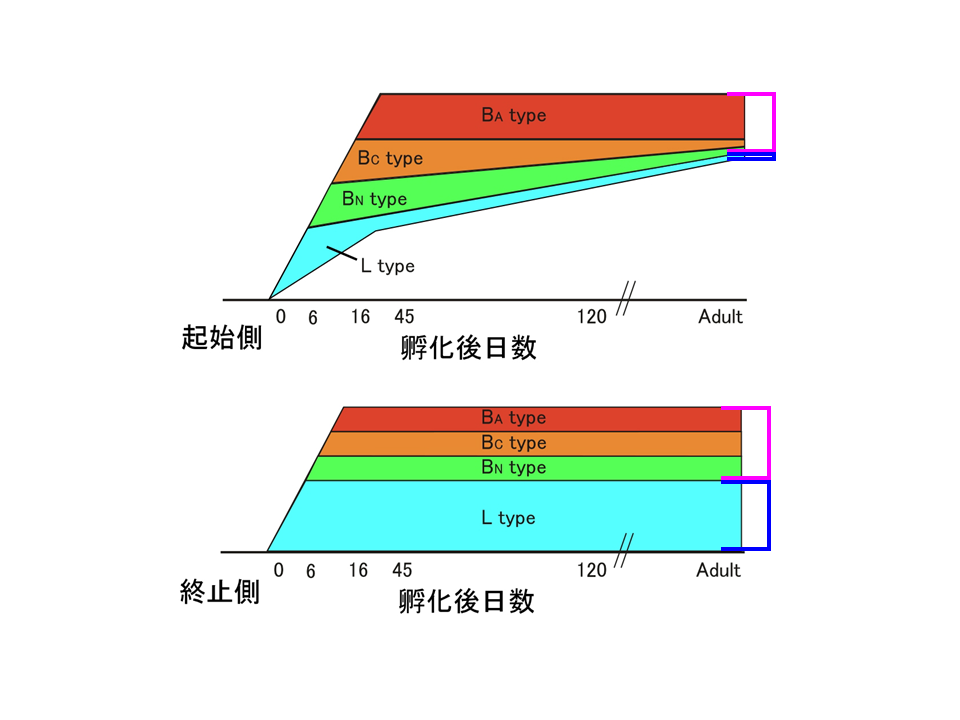
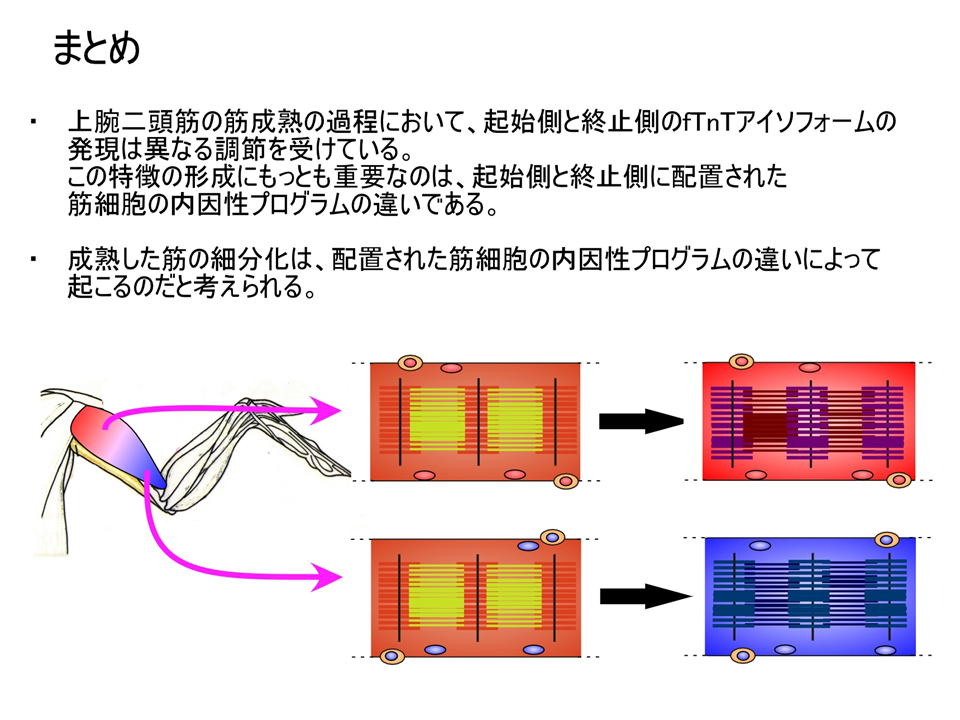
模式図で示すと、上腕二頭筋の肩側(起始側)では、最初にL型が多いが、しだいにB型に置き換わっていく。しかし、肘側(終止側)では、L型が減らず、B型(BN、,BC、BA)も孵化後6日目あたりでみられるようになり、その後は量的に大きな変化はない。つまり、上腕二頭筋は成熟の過程で、肩側(起始側)と肘側(終止側)で異なる調節を受けるようになる。

説明文の9
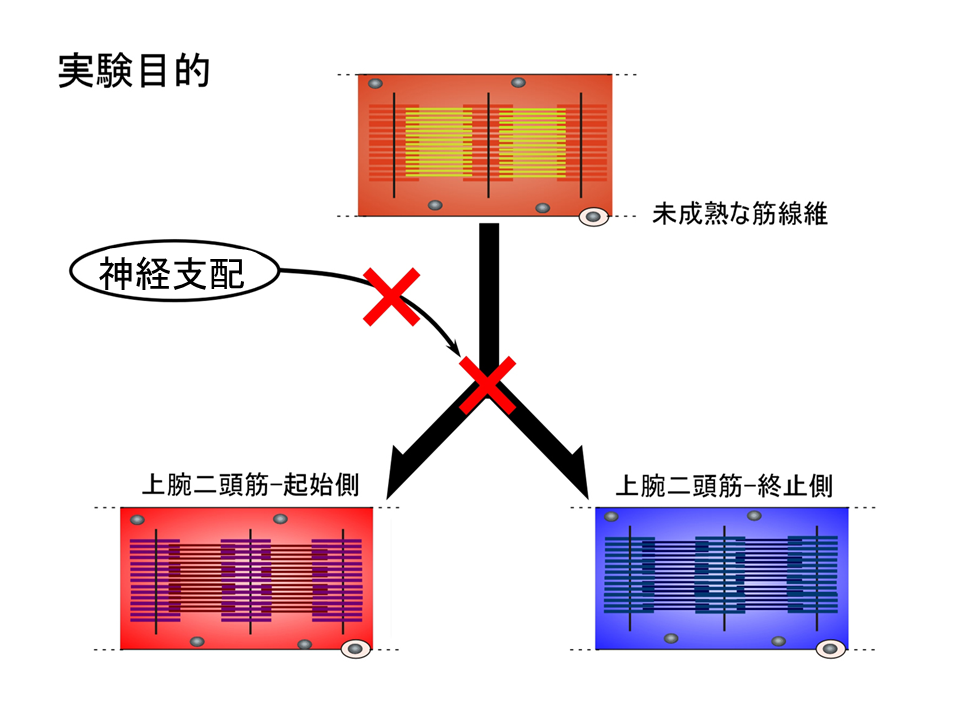
速筋や遅筋、混合筋への細分化を調べる前に、上腕二頭筋内の部位による違いへの神経支配の影響を調べることにした。
説明文の10
もし神経支配が筋肉の細分化の要因ならば、その神経支配を除神経操作により取り除いてしまえば、肩側(起始側)と肘側(終止側)の差異は形成されないはずである。孵化後1日目もしくは2日目の上腕二頭筋につながる正中神経を焼けたピンセットで切断し、除神経を行って、ある期間飼育した後、トロポニンTアイソフォームの発現パターンを調べた。いったん神経を切っても、また繋がってしまうことがあるが、手術したニワトリでは翼が常にだらんと垂れ下がっていたことから、神経は再連結していないことがわかった。
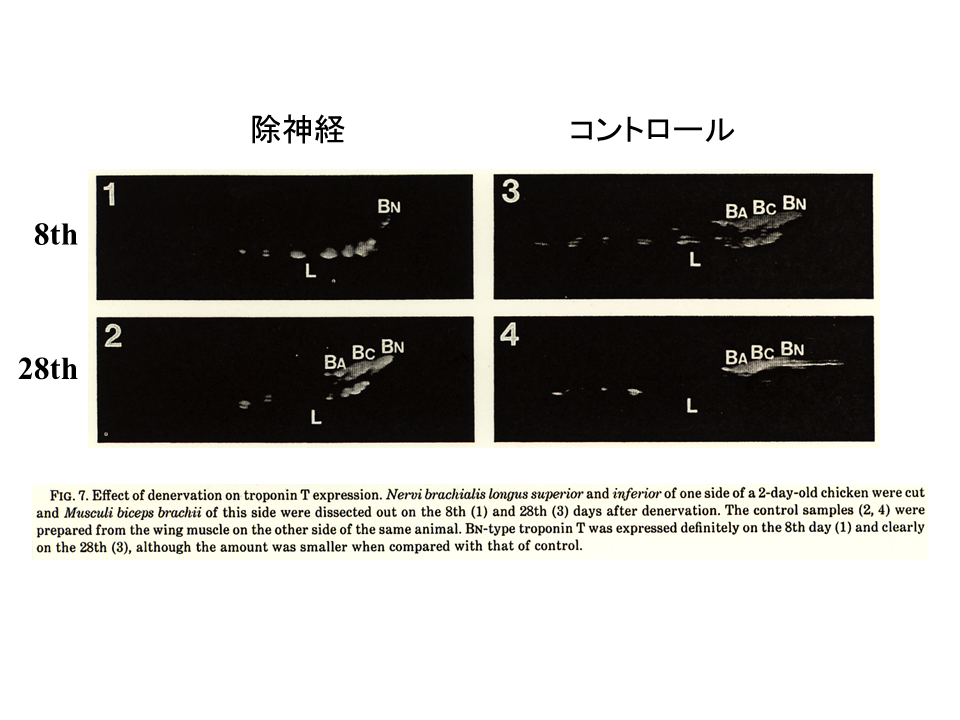
説明文の11
まず除神経手術の影響を調べた。片側の翼の神経を切断し、もう一方の翼の神経はそのままにした(コントロール)。手術によって発生が送れることがわかった。除神経してから28日目の上腕二頭筋のトロポニンTアイソフォームの発現パターンは、除神経しない側の8日目のパターンに似ている。すなわち、約20日間くらい発生が遅れたことになる。
説明文の12
除神経して30日目ぐらいから、肩側(起始側)と肘側(終止側)で違いがでてきた。手術をしているために、正常なものより変化が遅れるが、肩側(起始側)では、肘側(終止側)に比べて明らかにB型アイソフォームの発現量が多い。
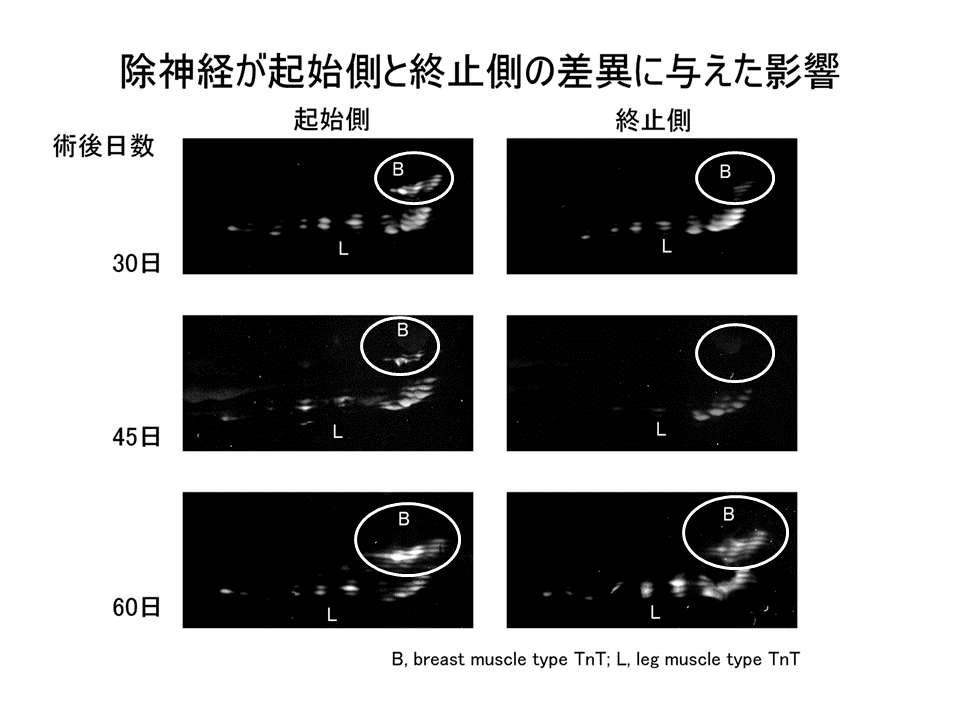
説明文の13
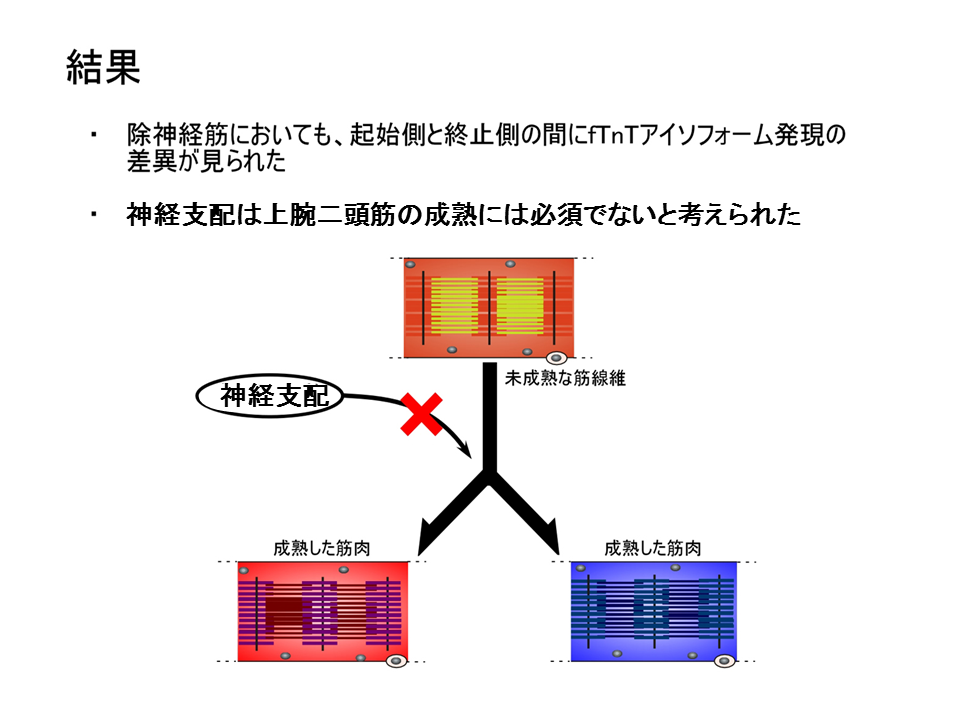
3回実験をおこなった結果、どの場合でも除神経した上腕二頭筋では、肩側(起始側)の方が肘側(終止側)よりB型に対するL型の量比(B/L)が大きかった。手術をしているため、コントロールと値が全く同じにはならなかったが、肩側(起始側)の方が肘側(終止側)よりB型に対するL型の量比(B/L)が大きいという点では全く一致した。すなわち、除神経しても肩側(起始側)と肘側(終止側)の差異が見られたことを示している。
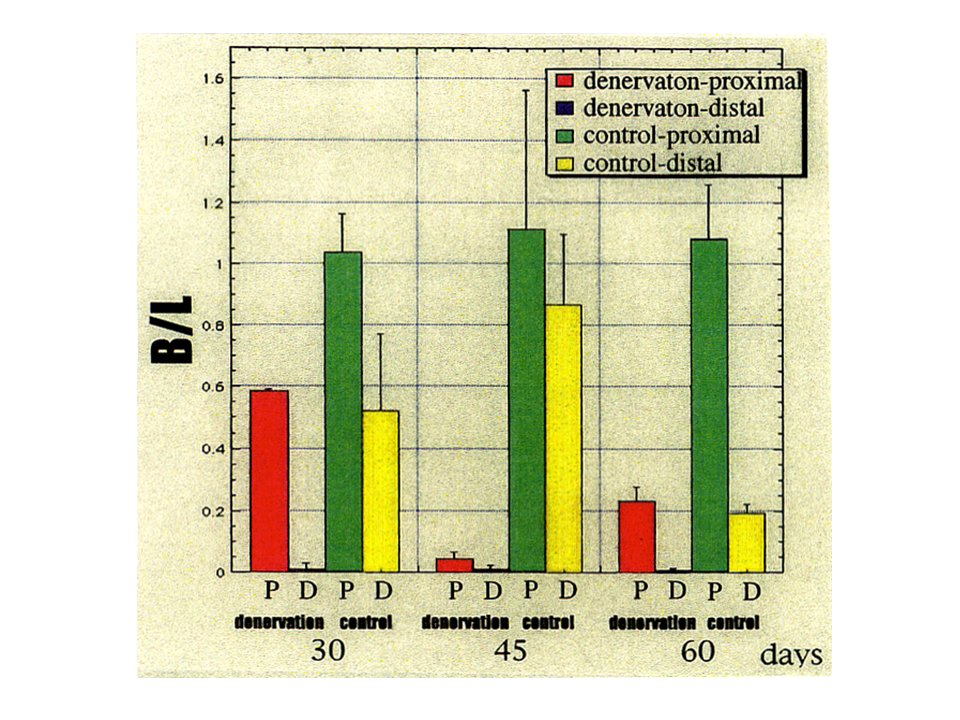
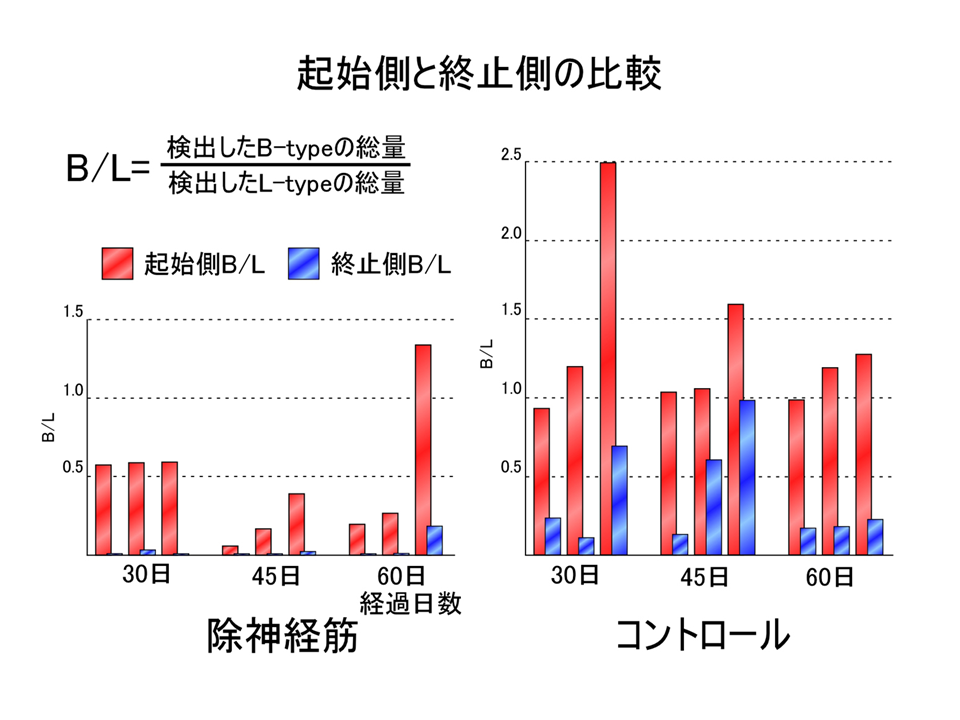
説明文の14
3回の実験結果の平均を示した。除神経した場合(赤色と青色で示す)、コントロール(緑色と黄色で示す)と全く同じにはならなかったが、肩側(起始側)の方と肘側(終止側)で部位による差異があらわれるという点では一致していた。
説明文の15
この結果から、除神経しても正常の場合と同じパターンが現れることがわかり、神経支配は上腕二頭筋の細分化に深く関わる要因ではないことが明らかとなった。
説明文の16
次に上腕に存在する位置情報が、細分化の要因となりうるかを調べた。
説明文の17
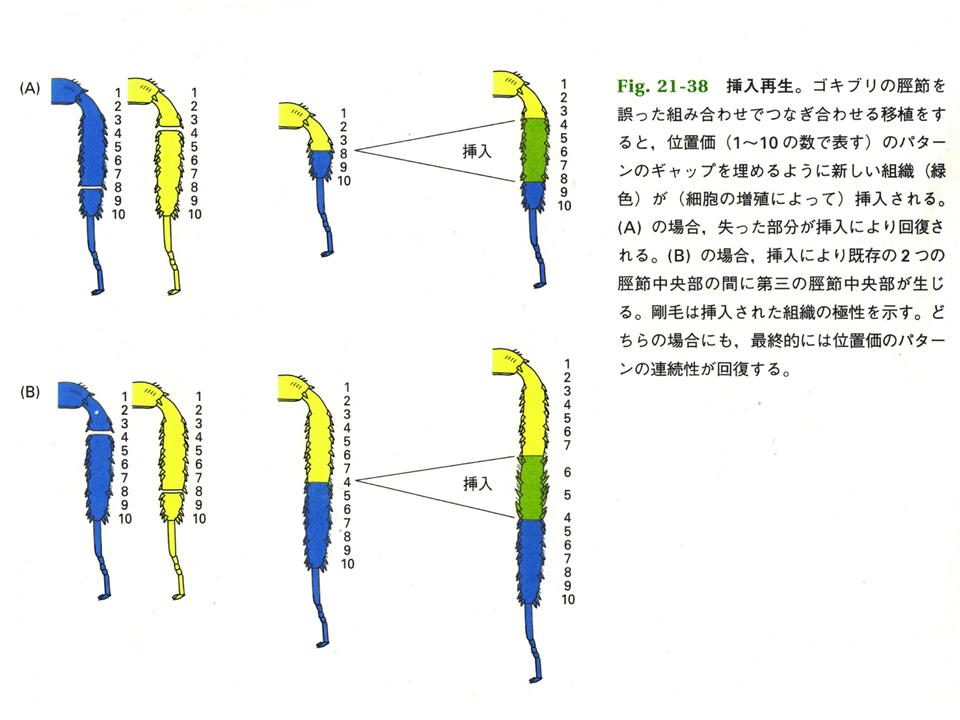
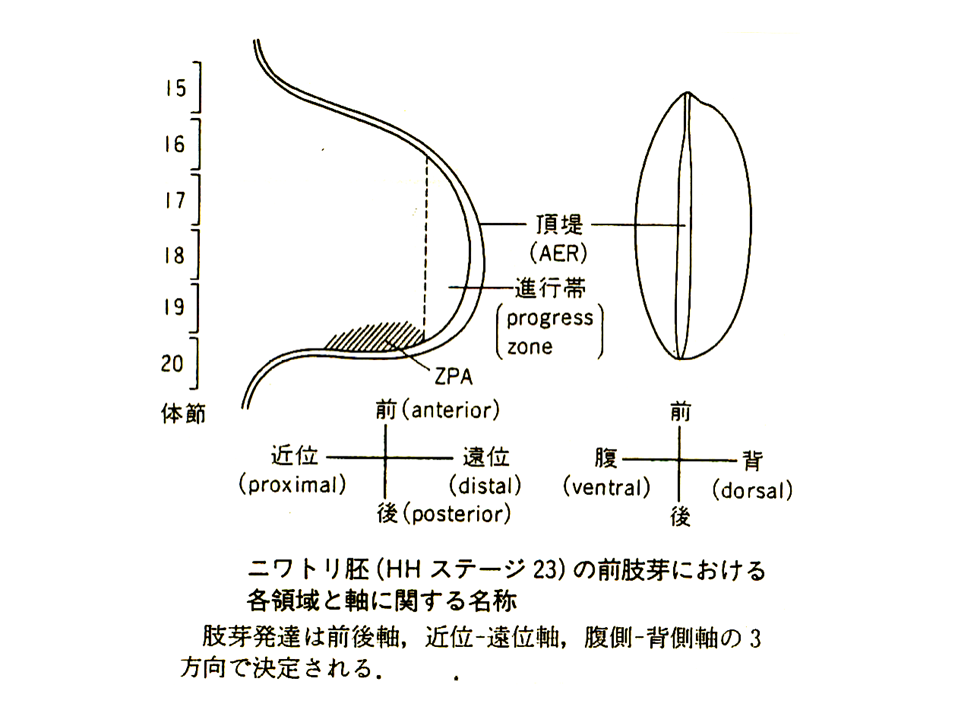
位置情報とは何かを示す面白い実験がある。黄色ゴキブリと青色ゴキブリの肢を切断し、黄色ゴキブリの肢の基部(1〜3)と青色ゴキブリの先端部(8〜10)を繋いでみると、その間の部分(4〜7)が再生した。ゴキブリの肢に1から10の位置情報があるとすると、その情報に従って再生したことになる。決して基部(1〜3)や先端部(8〜10)がその部分に再生することはない。さらに黄色ゴキブリの基部からもっと長い部分(1〜7)と青色ゴキブリの先端部かもっと長い部分(4〜10)を切断して繋いでみると、黄色ゴキブリの7と青色ゴキブリの4の間をうめるように、4〜6に相当する部分が再生した。位置情報に従って4〜6が再生したことになる。イモリの前肢を手首にあたる所から切断した場合、手にあたる部分は再生してくるが、決して腕に当たる部分が再生してくることはない、これも位置情報に従って構造が形成されることを示している。
説明文の18
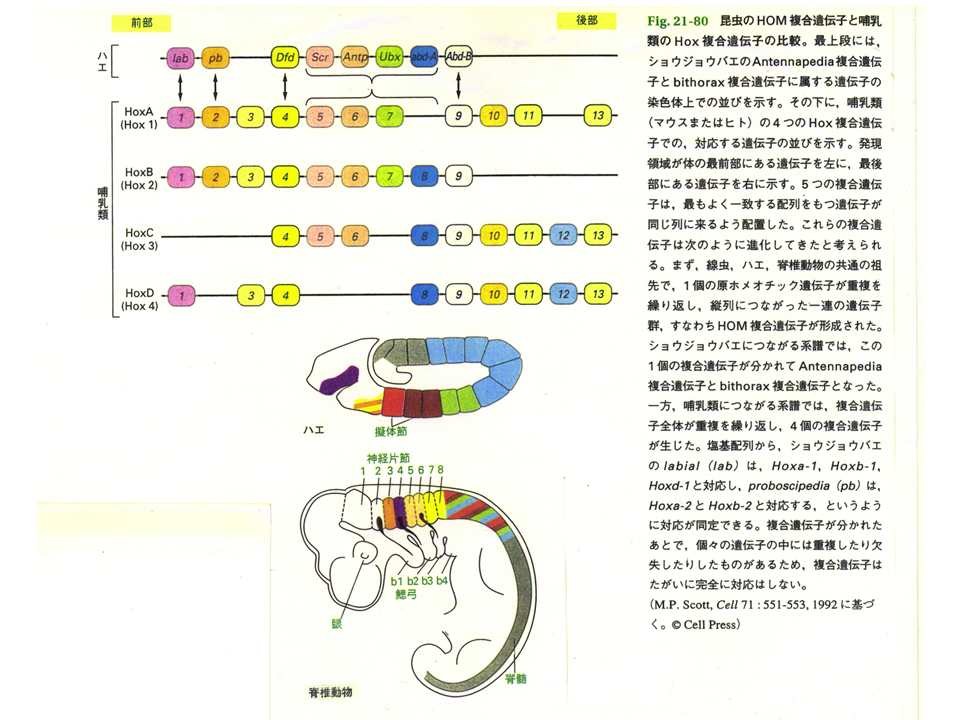
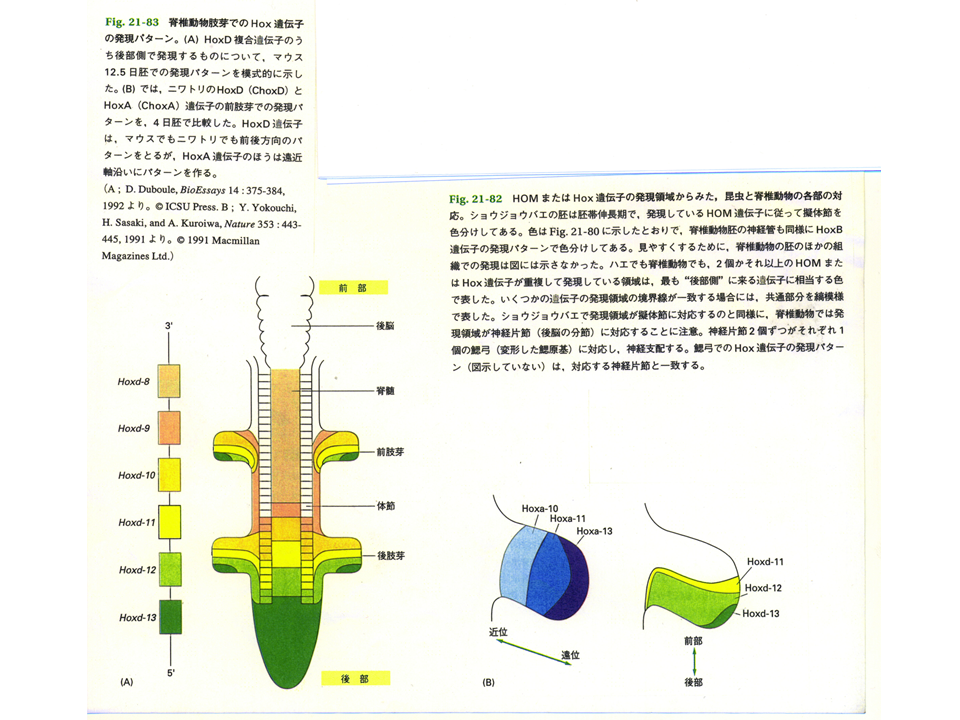
このような位置情報を作るものは何か?例えばショウジョウバエのHOM複合遺伝子群やほ乳類のHox複合遺伝子群は、遺伝子群を構成する各々の遺伝子が前後軸にそって順番に発現する。しかもその発現する順番どおりに遺伝子が同じ染色体上に並んでいる。すなわち、これらの複合遺伝子群が前後軸の位置情報を与えているとも言える。ショウジョウバエはHOM複合遺伝子群を1セットしかもたないが、ほ乳類はHox複合遺伝子群を4セットもっている。
説明文19
説明文の20
ほ乳類にはHox複合遺伝子群が4セット(Hoxa〜Hoxd)ある。Hoxdは前後軸に関係するが、なんとHoxaは近位遠位軸に沿って異なる遺伝子が発現しているのである。これが近位遠位軸に沿った位置情報を作り上げているかもしれない。
説明文の21
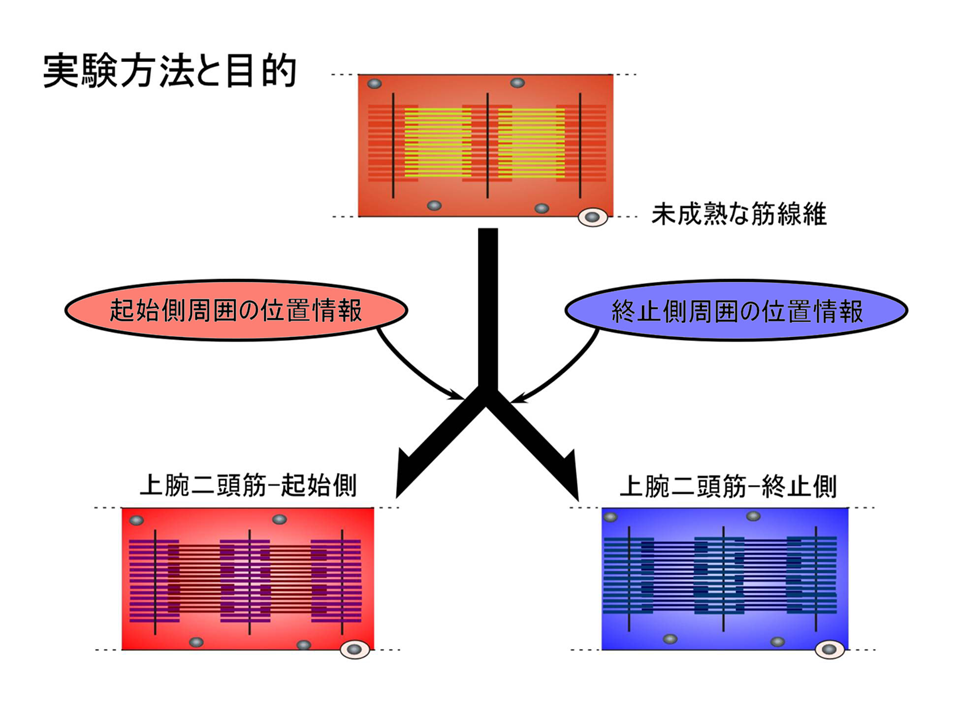
位置情報が細分化の要因だとすれば、その情報を乱した時にトロポニンTアイソフォームの部位における差異も乱れるはずである。
説明文の22
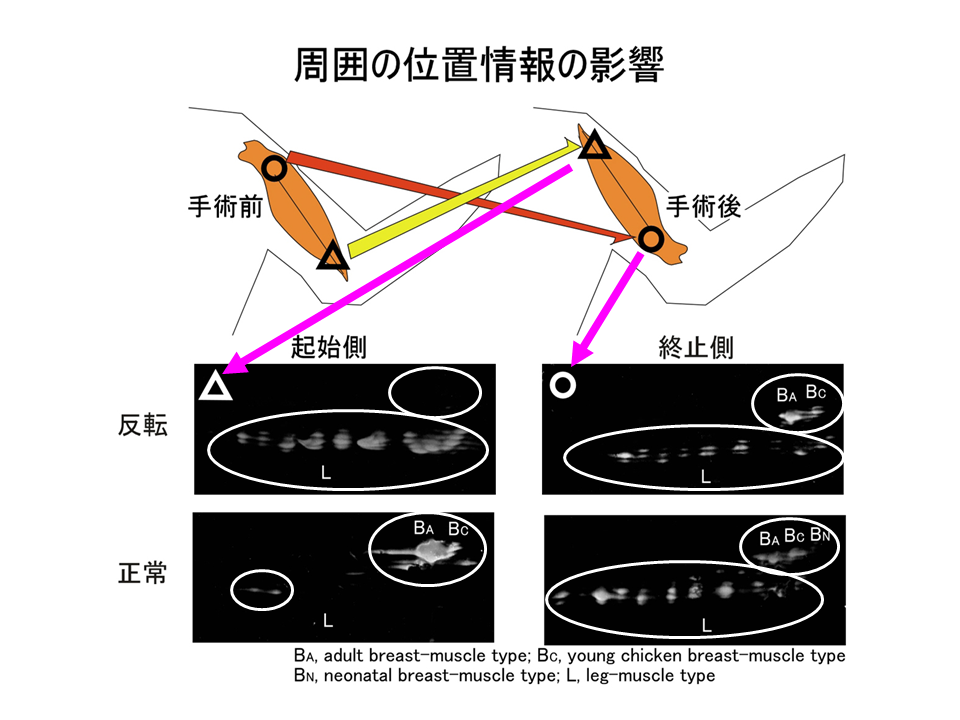
そこで、孵化後15日目のニワトリの上腕二頭筋を摘出し、肩側(起始側)の方と肘側(終止側)を反転させ、それぞれの腱を接着剤で固定し、120日間飼育した後、トロポニンTアイソフォームの発現パターンを調べた。もともと肩側(起始側)であり、反転後に肘側(終止側)となった部位では、もともと肘側(終止側)であり、反転後に肩側(起始側)となった部位よりも、B型に対するL型の量比(B/L)が大きかった。手術を行っているので正常の場合と完全には一致しないが、反転しても、しない場合と同様に肩側(起始側)では肘側(終止側)よりB/Lが大きくなるというパターンは変わらなかった。
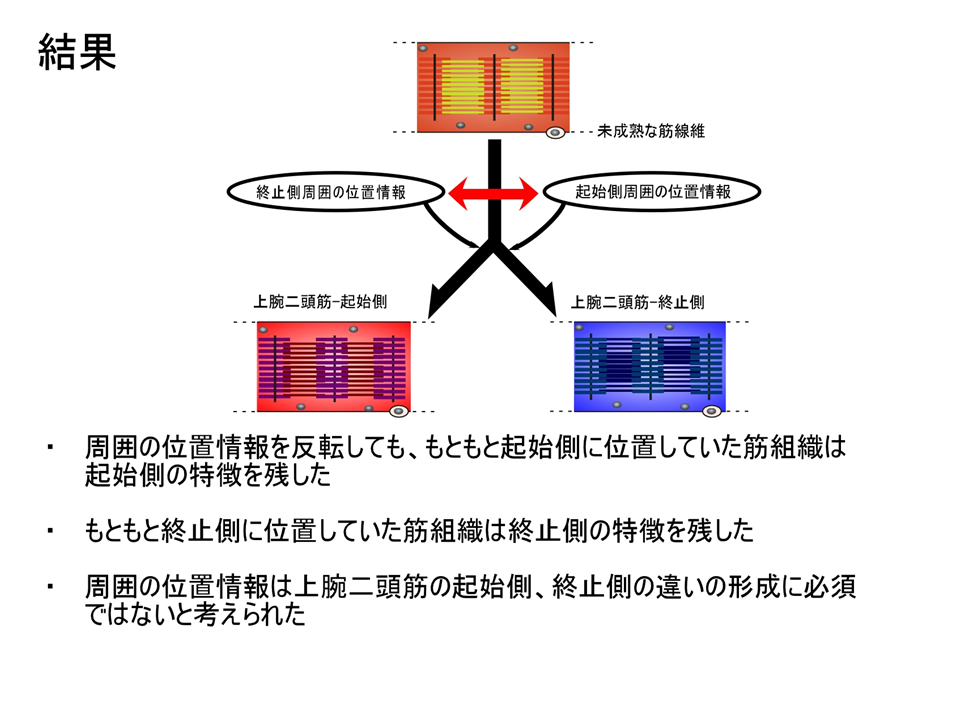
説明文の23
反転して位置情報を乱してもトロポニンTアイソフォームの部位的な差異は影響を受けないので、位置情報は上腕二頭筋の細分化に深く関わる要因ではないことが明らかとなった。ただし、実験を行った孵化後15日よりも前に位置情報がトロポニンTアイソフォームの発現パターンを決定してしまっており、その決定が反転しても覆されない場合、位置情報が関係ないとは言い切れなくなる。
説明文の24
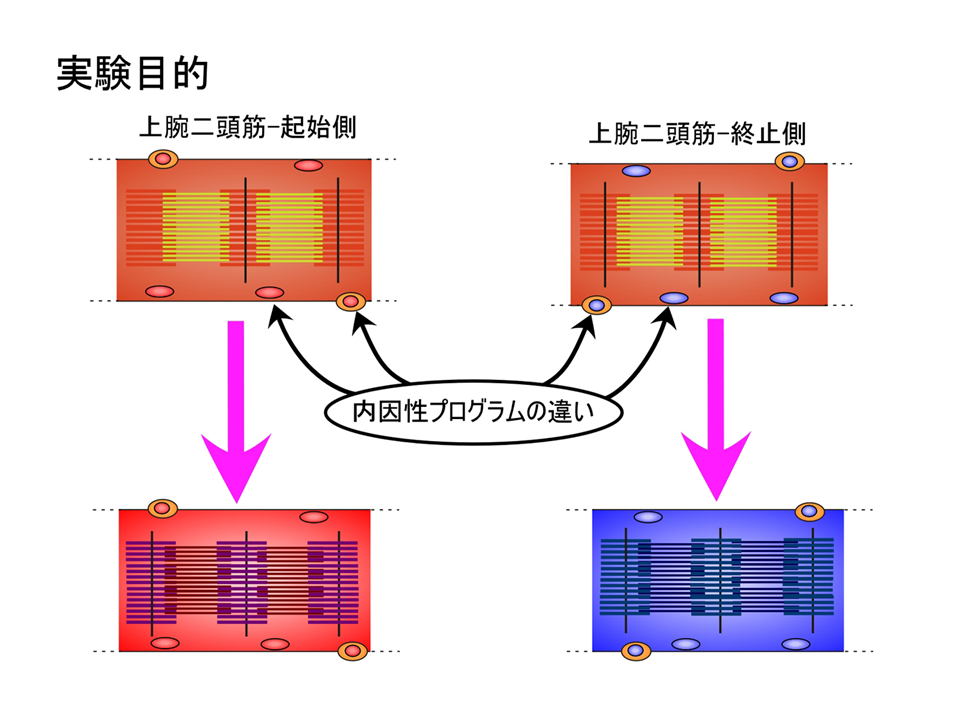
これまでホルモン、神経支配、周囲の位置情報がほぼ否定されたので、最後に内因性プログラムが上腕二頭筋の部位的な差異を生み出す要因であるか否かを調べた。
説明文の25
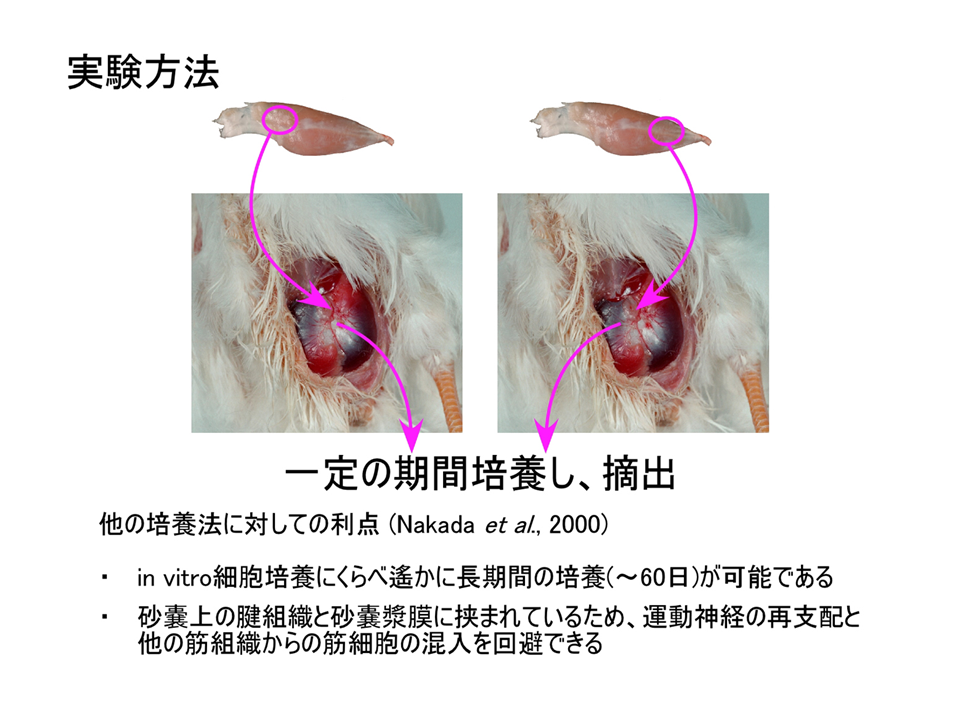
内因性プログラムの関与を調べるために砂嚢漿膜下組織培養法を利用した。砂嚢漿膜下には血管系が発達しており、それによって供給される酸素や栄養によって組織を培養することができる。砂嚢漿膜下には、筋肉を刺激する神経支配も、上肢にあるかもしれない位置情報も存在しない。上腕二頭筋の肩側(起始側)と肘側(終止側)を切断し、各々を砂嚢漿膜下に埋め込んで、長期間培養した後、トロポニンTアイソフォームの発現パターンを調べた。
説明文の26
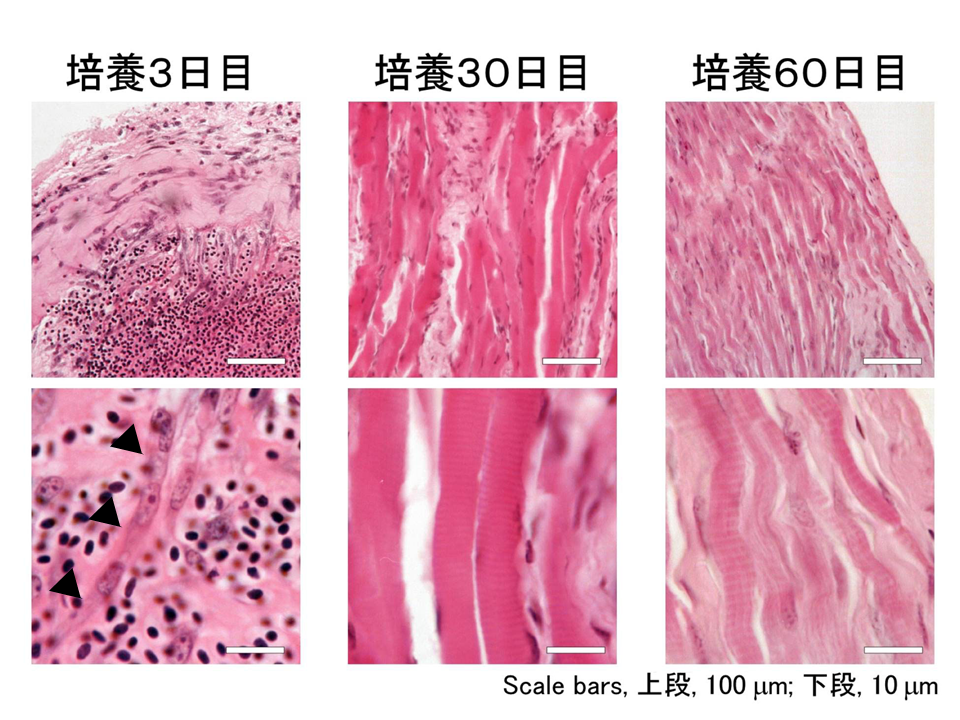
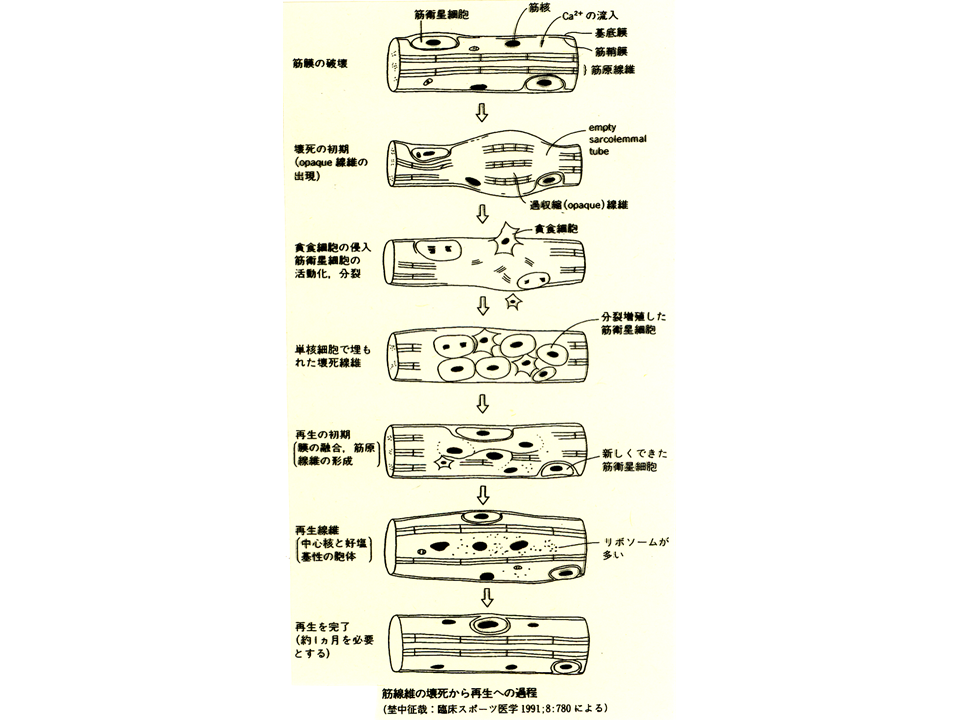
砂嚢漿膜下で培養された筋組織は、移植後3日目で一度ほぼ完全に変性(殆どの細胞が死んで構造が壊れること)し、その後に衛星細胞が分裂・融合して筋組織が再構築された。スケールバーは50マイクロメートル。
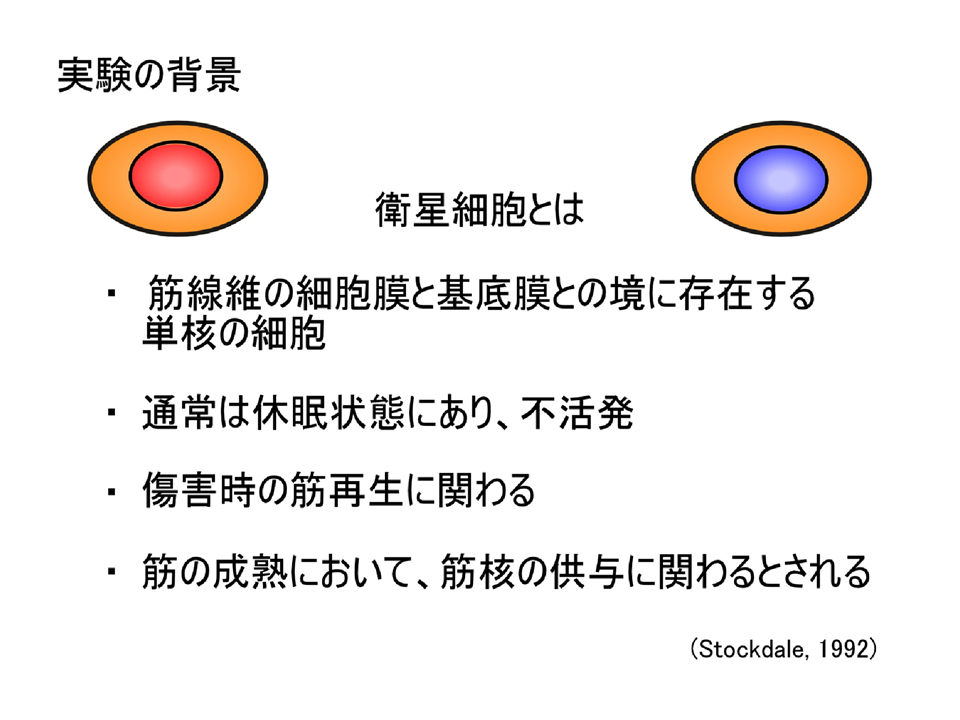
説明文の27
衛星細胞とは、筋線維(筋繊維)の細胞膜と規定膜の間に存在し、分裂をせず休眠状態にある細胞である。
説明文の28
衛生細胞は筋が傷害を受けると分裂・融合して筋を修復する。
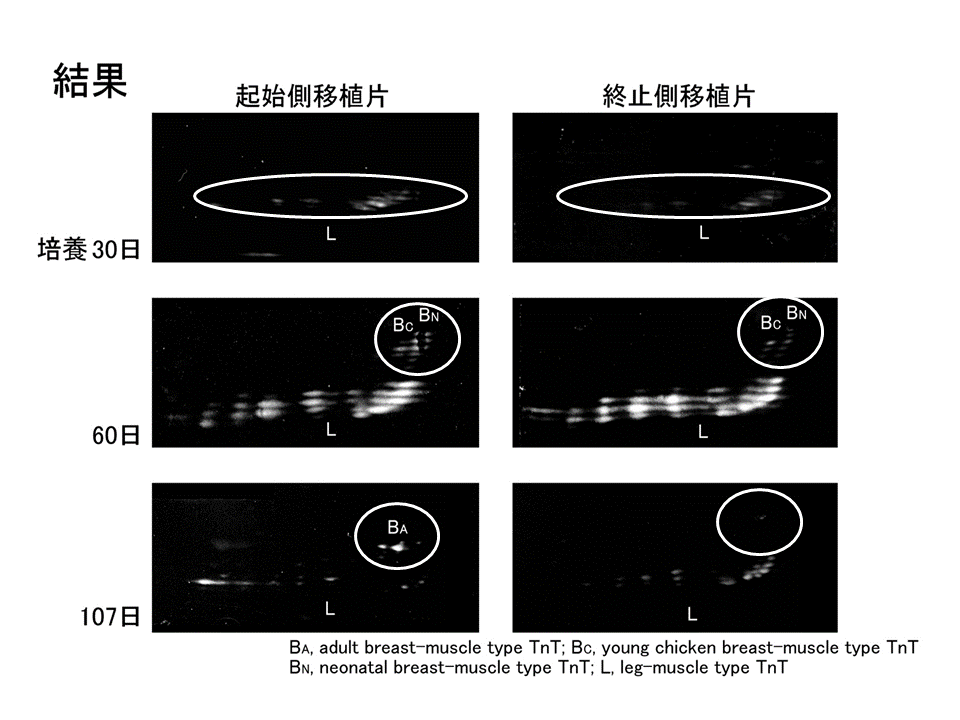
説明文の29
砂嚢漿膜下で肩側(起始側)部分と肘側(終止側)部分を各々培養した。培養30日では、再生筋線維が未熟な状態にあり、L型だけが発現しており、肩側(起始側)の方と肘側(終止側)で違いがみられなかった。培養期間が長くなると、肩側(起始側)では肘側(終止側)に比べB型の発現量が多くなった。
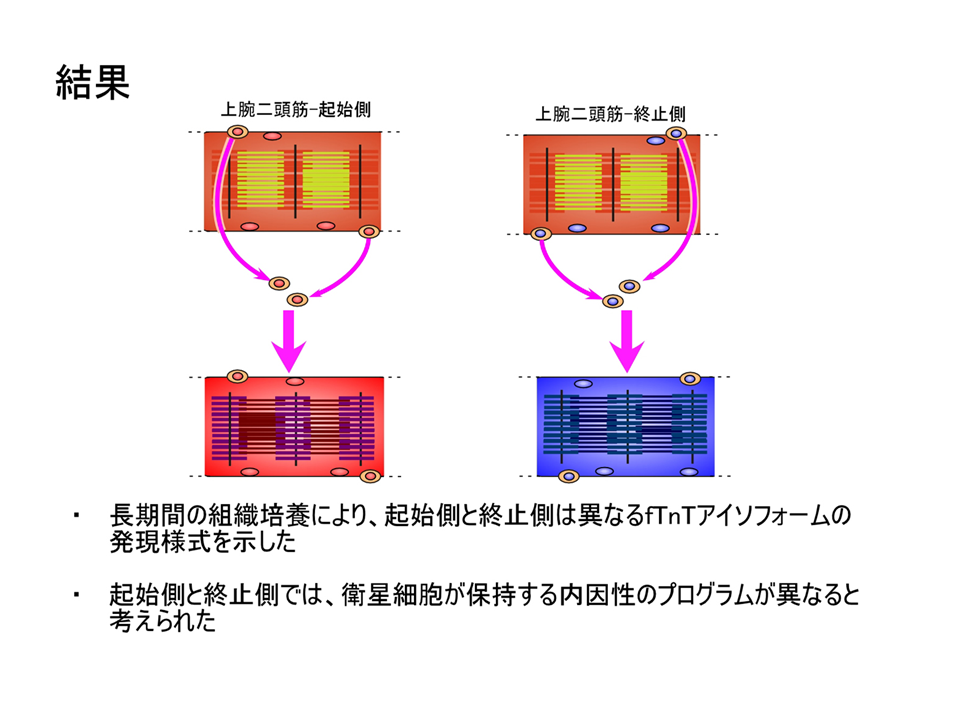
説明文の30
神経支配も位置情報も存在しない、砂嚢漿膜下での長期間の組織培養によって、肩側(起始側)では肘側(終止側)よりB型に対するL型の量比(B/L)が大きくなった。手術、培養を行っているため、正常発生と全く同じではないが、正常発生に似たパターンが得られた。
説明文の31
これらの結果から、上腕二頭筋における、トロポニンTアイソフォームの発現パターンは、ホルモン、神経支配、周囲の位置情報によって決められるのではなく、内因性プログラムによって決められることが明らかとなった。それでは内因性プログラムとはいったい何なのか?それはまだわからない。しかし、筋肉の発生をみると、中胚葉細胞の中で、MyoDあるいはMyf5という遺伝子が働くと、その細胞は筋肉に成るべく運命が決まり、さらにmyogeninという遺伝子が働くと、運命に従って実際に筋肉になっていく。つまり、遺伝子の発現によって筋発生は制御されている。それゆえ細分化も遺伝子の発現によるのではないか。myogeninが働いた後、何らかの遺伝子か遺伝子群が働いて細分化され、それに基づいて筋肉の収縮・弛緩を調節する遺伝子群(例えばトロポニンT)が働くのではないだろうか。
説明文の32
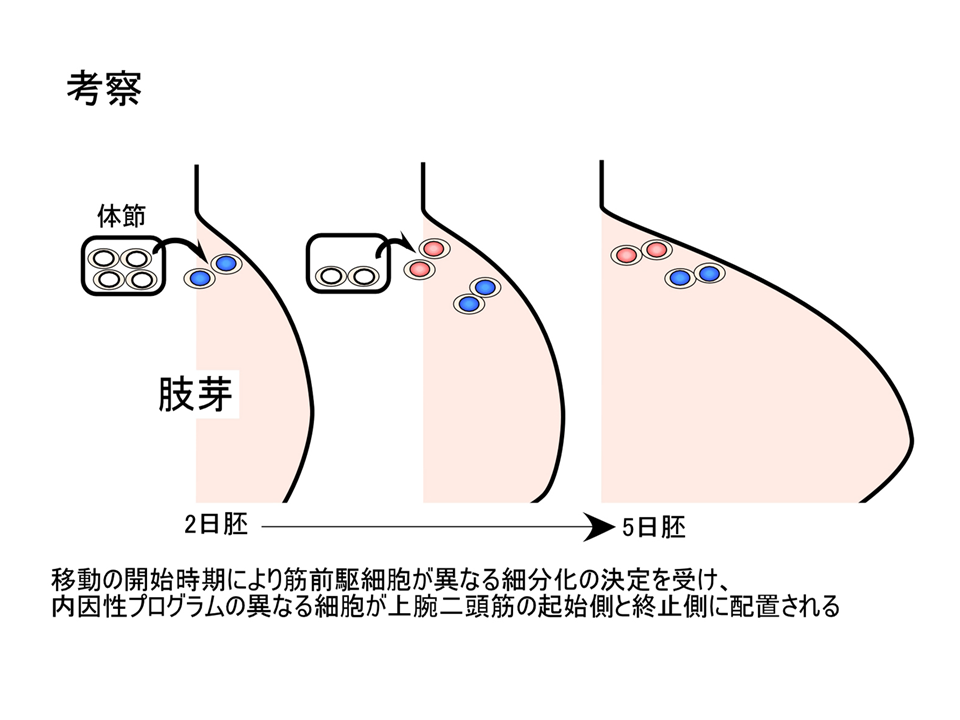
その内因性プログラム(遺伝子発現)はいつ、どこで決定されるのであろうか。ニワトリの上肢の筋肉のもとは、始めからそこにあるのではなく、体の中央にある体節から移動して来る。MyoDあるいはMyf5による運命を決める決定と、myogeninによる運命に従って実際に筋肉を形成する分化にタイムラグがあるため、体節から上肢へ筋肉のもとが移動できる。タイムラグがなかったらおそらく筋肉のもとは体節周辺で筋肉になってしまう。肩側(起始側)と肘側(終止側)でトロポニンTアイソフォームの発現パターンが異なるのは、筋肉のもとが移動してく時期の差かもしれない。最初に移動してきたものはより奥深くに入り込んで肘側(終止側)の内因性プログラムをもつようになり、後から移動してくるものはそれほど入り込むことはなく肩側(起始側)の内因性プログラムをもつことになる。あるいは体節内で時期によって違ったプログラムが組まれるのかもしれない
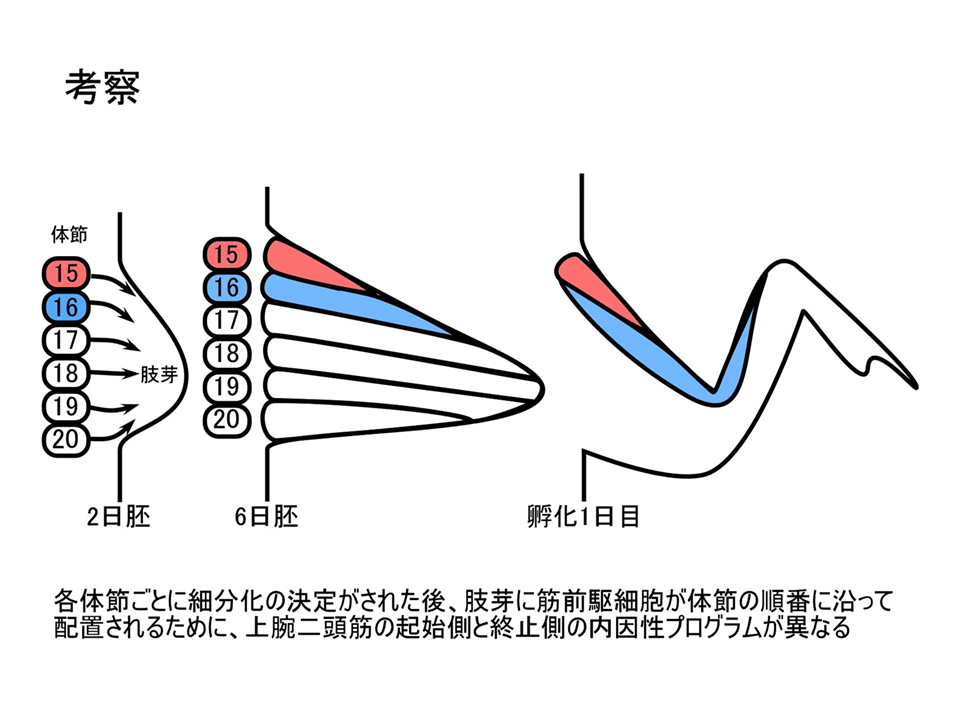
説明文の33
また、異なる体節で異なるプログラムが組まれるのかもしれない。例えば体節15では肩側(起始側)のプログラム、体節16では肘側(終止側)のプログラムといったように。
説明文の34
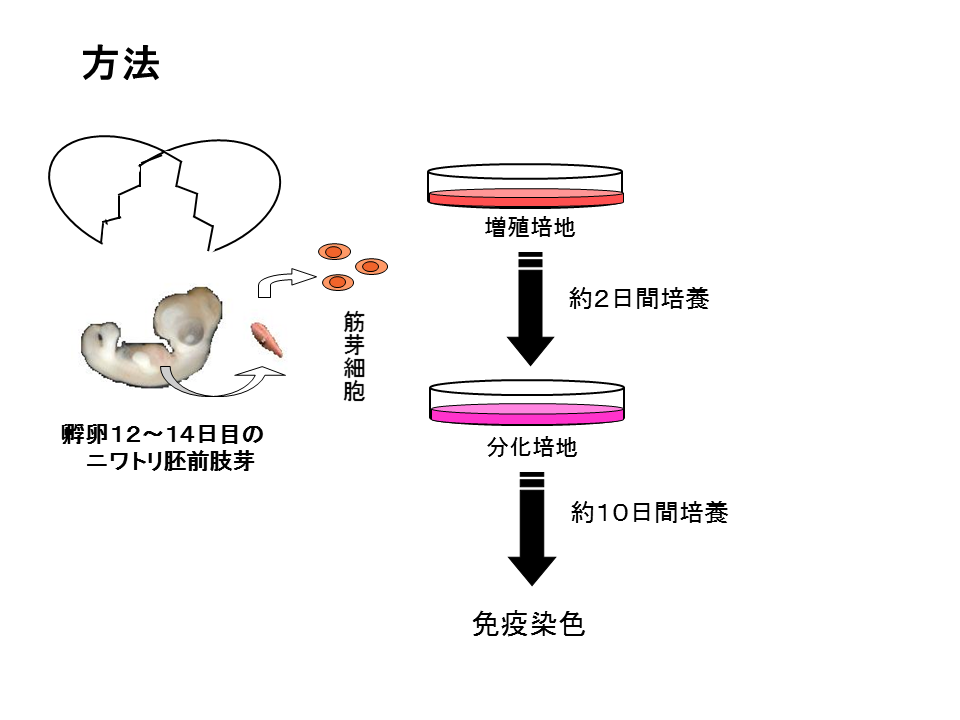
時期によって違うプログラムが組まれる可能性を調べてみた、ニワトリの孵化前の12〜14日目の胚から、将来下肢となる肢芽を切断し、半分(基部側と先端側)に切って細胞培養してみた。筋繊維はいったん変性して、その後衛星細胞が新たな繊維を構築する。

説明文の35
13日胚。新たに形成された筋繊維(左上)を、B型トロポニンTだけと反応する抗体を用いて調べた。この抗体の結合した繊維は赤く見える。基部側(起始側)では線維全体でB型が発現しており、さらに局所的に強い発現が見られた。
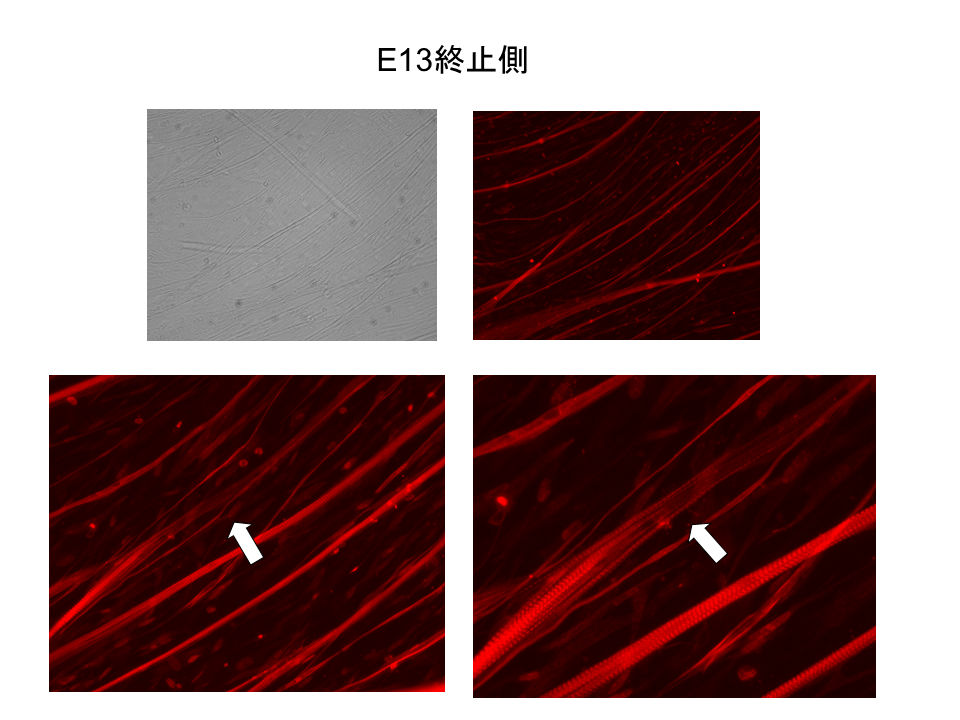
説明文の36
一方、末端側(終止側)では、線維全体でB型の発現がみられることはほとんどなく、 局部的に弱い発現が見られた。

説明文の37
14日胚。13日胚と同様に、基部側(起始側)では線維全体でB型が発現しており、さらに局所的に強い発現が見られた
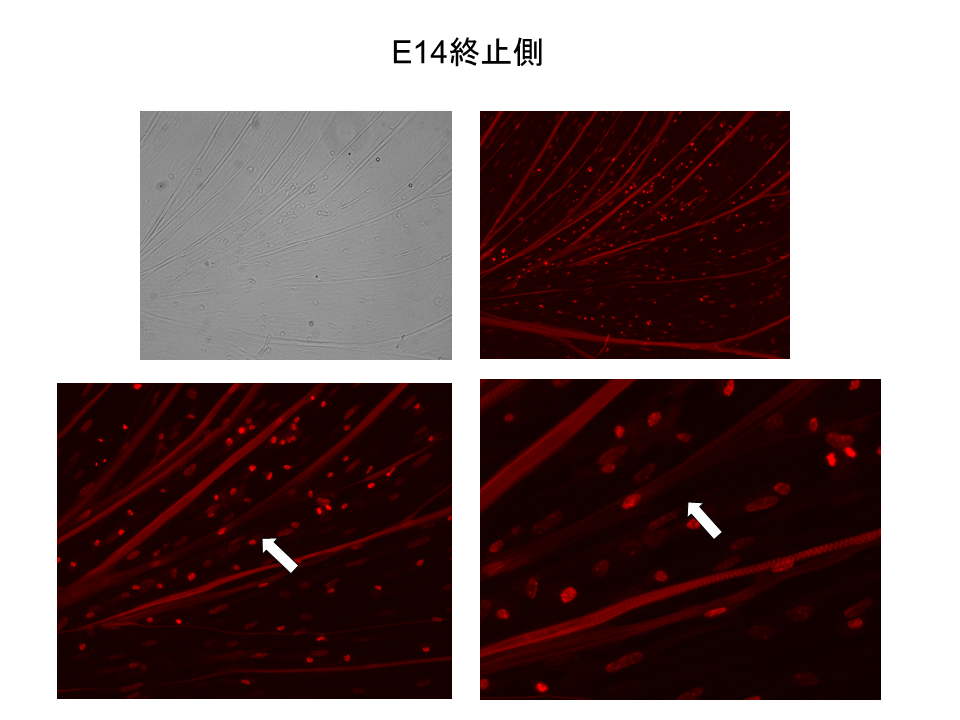
説明文の38
13日胚と同様に、末端側(終止側)では、線維全体でB型の発現がみられることはほとんどなく、 局部的に弱い発現が見られた。
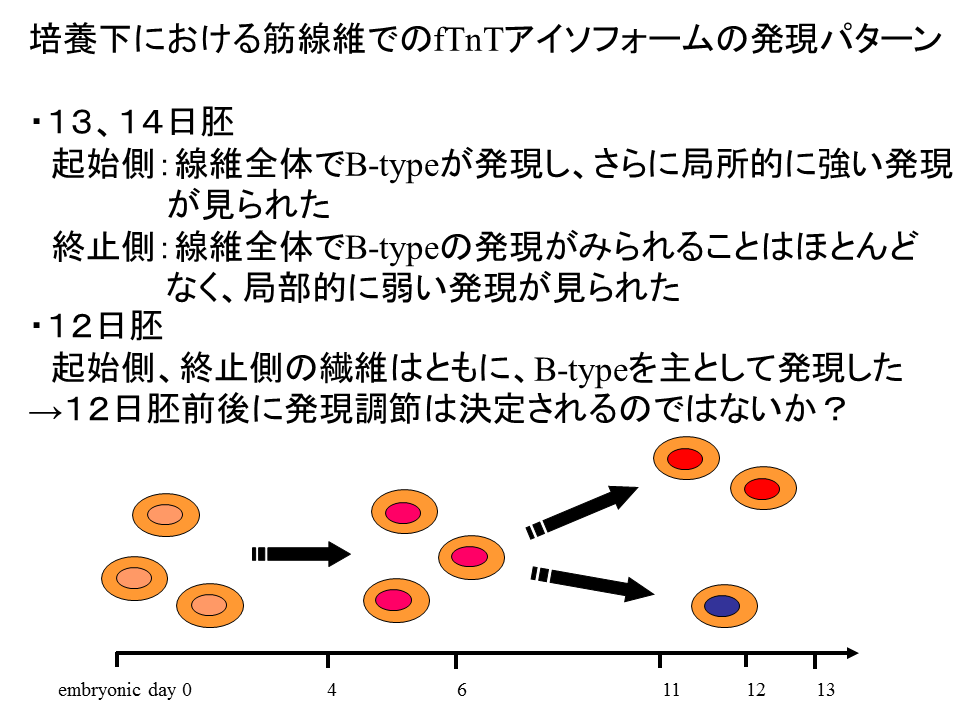
説明文の39
12日胚では、基部側(起始側)でも末端側(終止側)でもB型を主として発現がみられた。したがって、12日胚前後に発現調節の違いが決定されるのではないか?これらの点についてはさらに実験を重ねて、明らかにしていかなければならない。