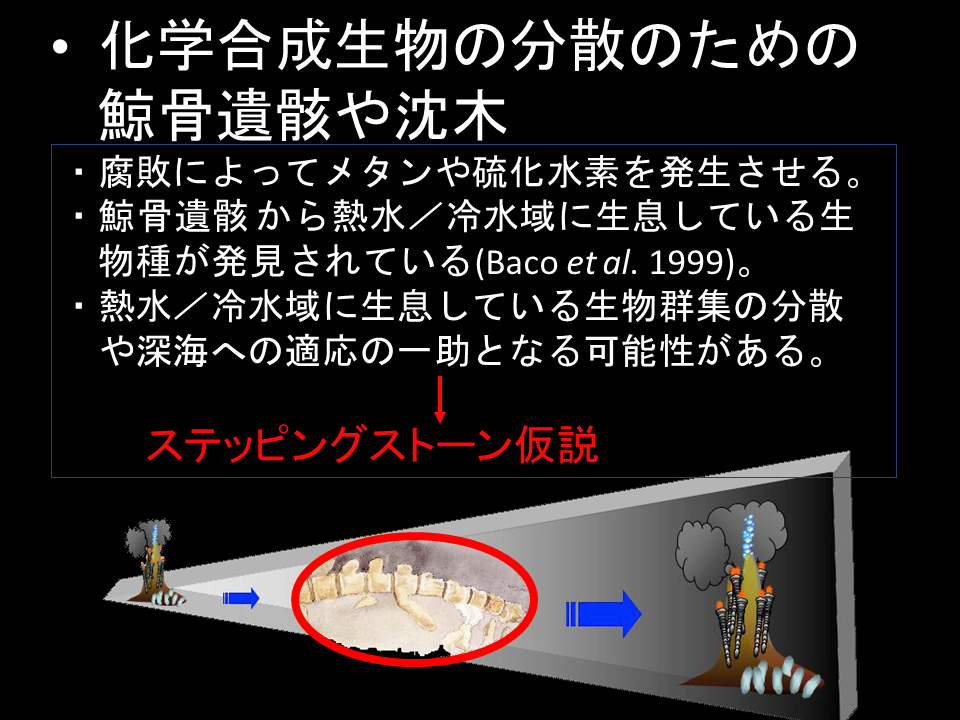
説明文の1
熱水域や湧水域の生物は、鯨遺骸や沈木を経由して分散するとする「地理的ステッピングストーン説」が提唱されている。しかし、中期地点があるにせよ、高い分散能力は必要であると思われる。
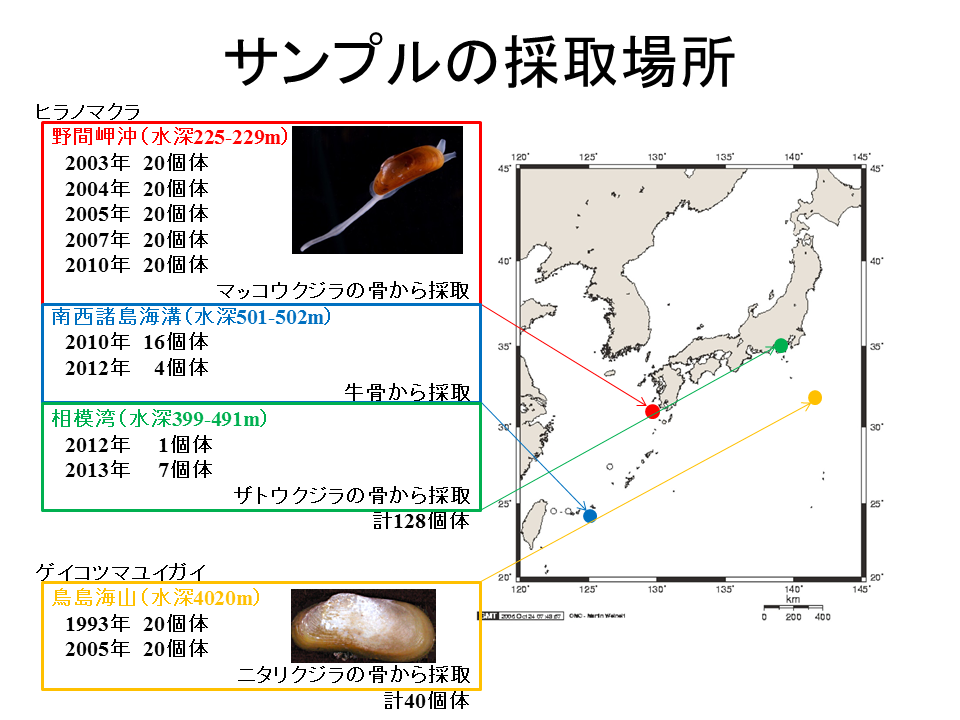
説明文の2
これはサンプルの採取場所を示した。ヒラノマクラは、野間岬沖では2003年から2010年の各20個体(計100個体)、南西諸島海溝では20個体、相模湾では8個体採集された。ゲイコツマユイガイは、鳥島海山で1993年と2005年に各20個体(計40個体)採集された。SmithとBaco(2003)は、コククジラ(Eschrichtius robustus)の鯨骨遺骸が、平均100km2に1体の割合で海底に存在すると推測しているが、 日本近海ではそれほどの頻度で発見されておらず、鳥島海山のニタリクジラしか自然死した鯨は見つかっていない。このように、自然死した鯨を捜し出すことは非常に難しいため、人工的に設置された鯨骨遺骸及び牛骨からヒラノマクラが採集された。
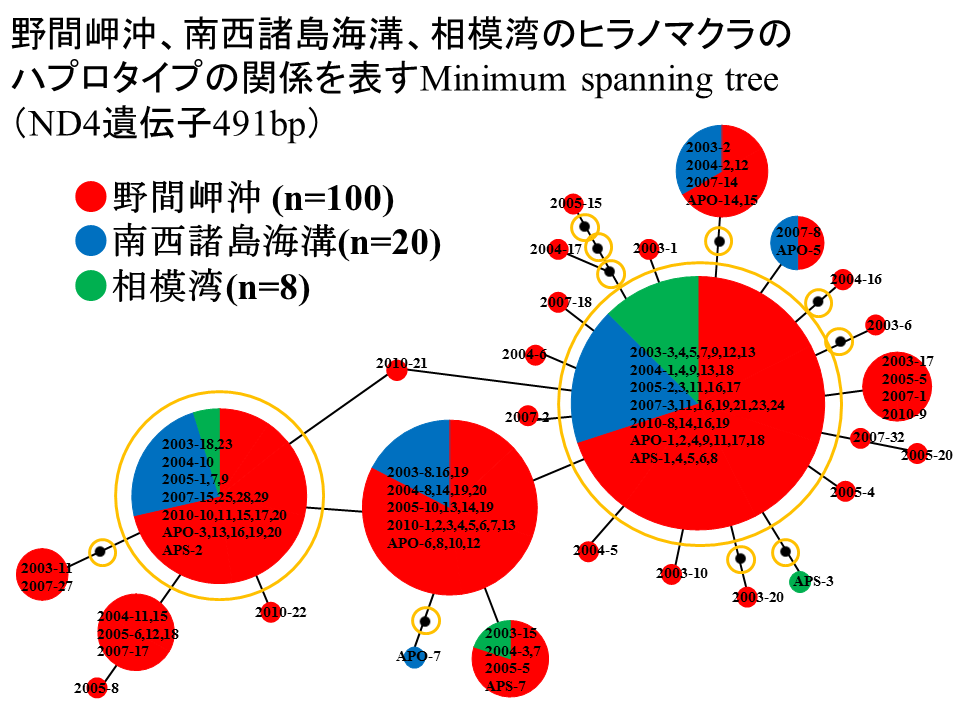
説明文の3
遺伝的な構成を調べるために、2003年から2010年の野間岬沖のサンプルを1グループ、2010年と2012年の南西諸島海溝のサンプルを1グループ、2012年と2013年の相模湾のサンプルを1グループとして、ND4遺伝子491bpに基づくミニマムスパニングツリーを構築した。野間岬沖を赤色、南西諸島海溝を青色、相模湾を緑色で示す。ミニマムスパニングツリーを構成している円はハプロタイプを表しており、面積がそのハプロタイプをもつサンプルの数に比例する。共通のハプロタイプをもつサンプルが3地点から発見されるなど、野間岬沖、南西諸島海溝、相模湾の3地点において、ハプロタイプの共通性が高いことがわかった。
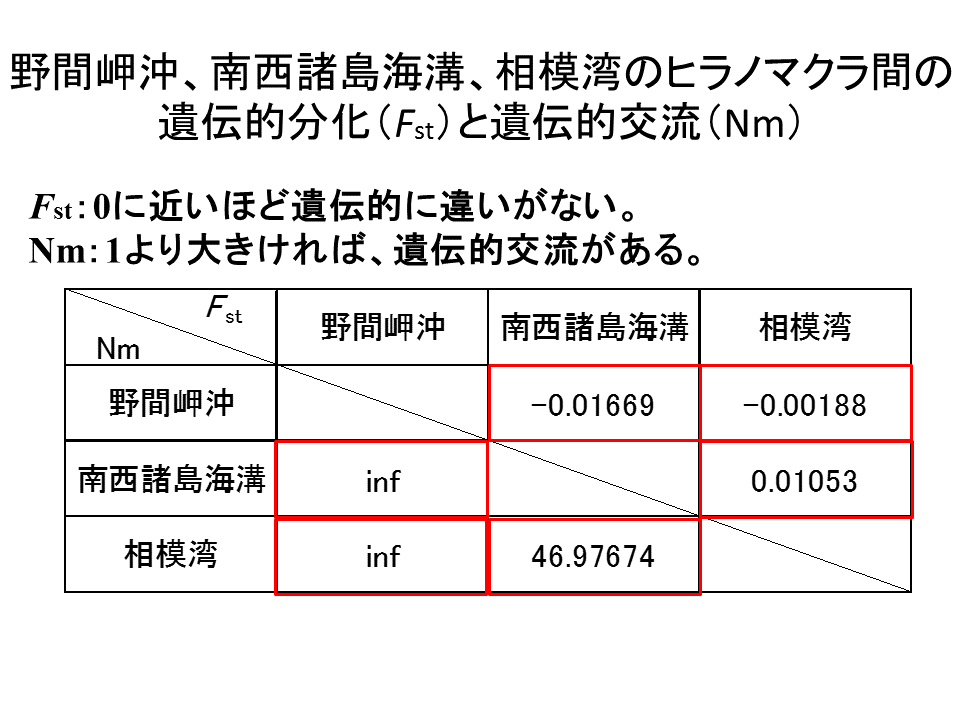
説明文の4
全ての地点間で、Fstの値はほぼ0と見なせる値となった。野間岬沖-南西諸島間、野間岬沖-相模湾間でNmの値は無限大となり、南西諸島海溝-相模湾間でも、1を遥かに越える値となった。

説明文の5
野間岬沖、南西諸島海溝、相模湾で多くのハプロタイプが共有され、また、3地点間の遺伝的分化が小さいことが示された。これらから、ヒラノマクラには地理的に隔たりのある3地点間での遺伝的交流が可能なだけの、非常に高い分散能力が備わっていることが明らかとなった。高い分散能力は地理的ステッピングストーン仮説が成立するための前提条件であり、ヒラノマクラで地理的ステッピングストーン仮説が成立する可能性が示唆された。
説明文の6
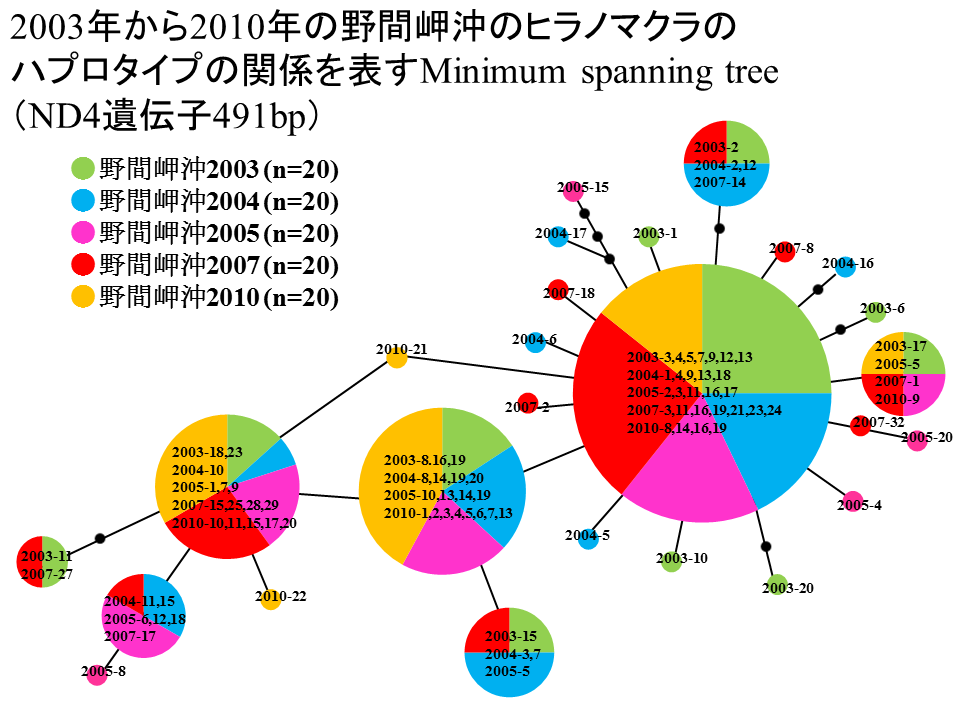
野間岬沖では2003年から2010年の間に豊富にヒラノマクラを採取することができたので、個体群の経年変化を調べた。2003年から2010年の野間岬沖のヒラノマクラを年度ごとに1グループとして、ND4遺伝子491bpに基づくミニマムスパニングツリーを構築した。2003年を緑色、2004年を青色、2005年を桃色、2007年を赤色、2010年を黄色で示した。2003年から2010年の間で、ハプロタイプの共通性が高く、大きく異なるハプロタイプ組成をもつ年はなかった。
説明文の7
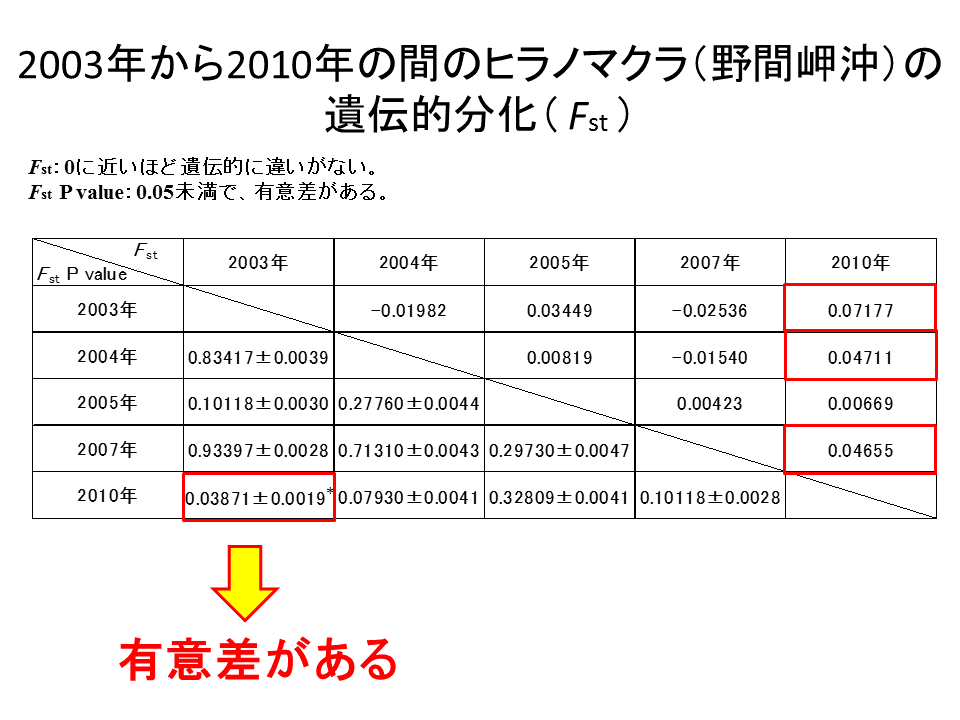
2003年から2010年の間のヒラノマクラのFst及びFst P valueの値を算出した。全ての年の間で、Fstの値はほぼ0と見なせる値となったが、2010年を含む組み合わせでは比較的大きいFstの値を取る傾向にあった。2003年-2010年間のFst P valueは、0.05を下回り、有意差があるという結果が得られた。
説明文の8
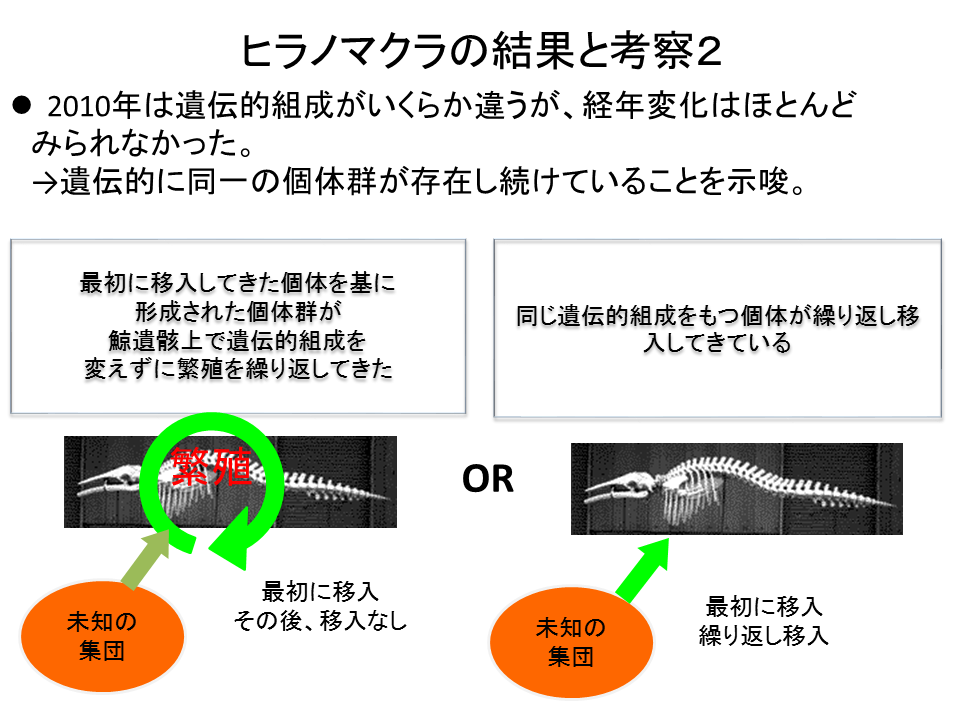
2010年は遺伝的組成がいくらか違うものの、野間岬沖に経年変化はほとんどみられなかった。このことは、遺伝的に同一の個体群が鯨骨遺骸上に存在し続けていることを示している。これは、最初に移入してきた個体を元に形成された個体群が鯨骨遺骸上で繁殖を繰り返してきたか、もしくは安定した規模の大きな未知の集団から、同じ遺伝的組成を持つ個体が繰り返し移入してきたものと推定される。後者が起こっていたとすれば、それは分散能力が高いためであると言える。

説明文の9
2003年から2007年で、Haplotype diversityは約0.86-0.90の値を取り近似したが、2010年では比較的低い値となった。2003年から2007年のNucleotide diversityは近似したが、2010年では半分程度の値となった。鯨骨遺骸の鯨油が消費され、硫化水素の発生量が少なくなったことで個体数が減少し、それに伴って遺伝的多様性も小さくなった可能性がある。
説明文の10
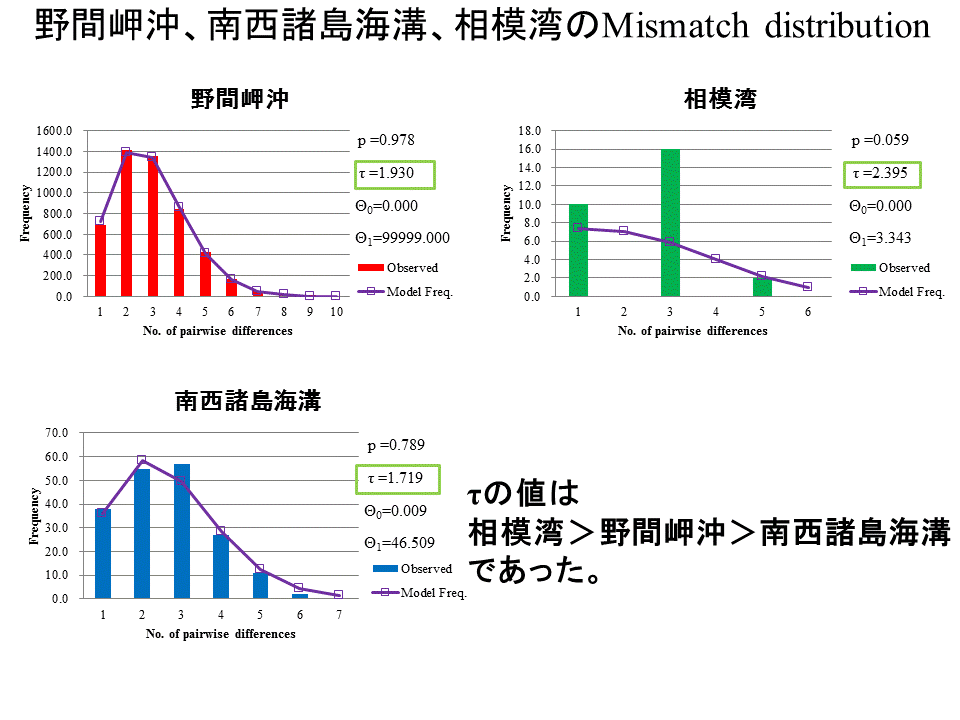
野間岬沖、南西諸島海溝、相模湾で集団が確立した順番を調べるため、3地点のMismatch distributionを算出した。横軸はハプロタイプ間で異なる塩基数を表し、縦軸はそれがどの程度の頻度で現れるかを表す。棒は実測値で、曲線はモデル値を示す。集団が確立してからの相対的な関係を表すτの値に注目すると、τの値は相模湾が一番大きく、次いで野間岬沖、南西諸島海溝が一番小さいという結果になった。
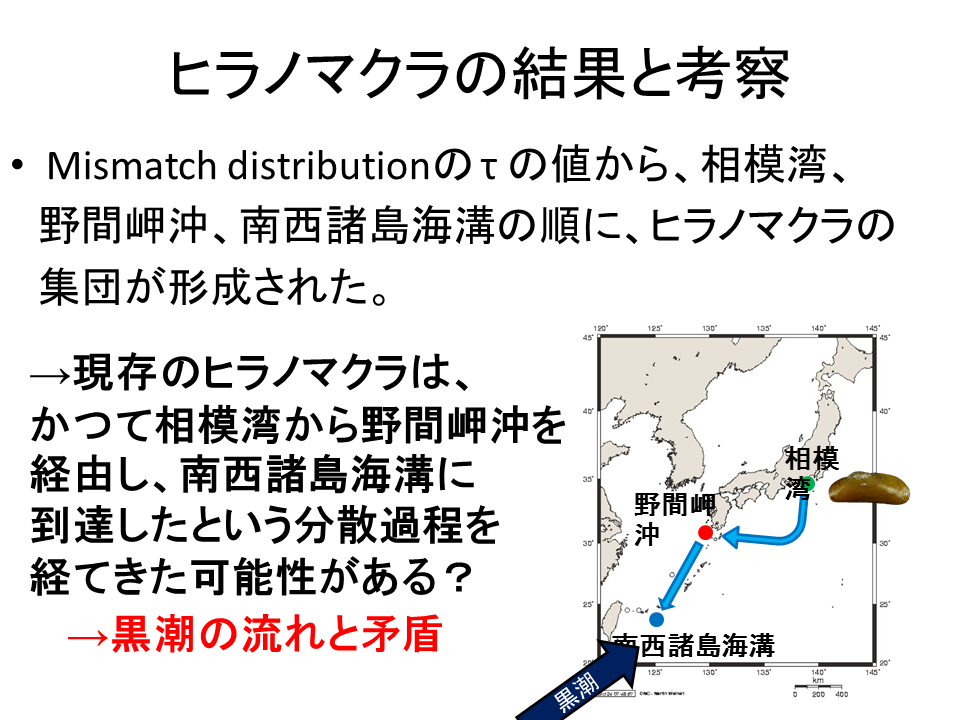
説明文の11
Mismatch distributionの τ の値から、相模湾、野間岬沖、南西諸島海溝の順に、ヒラノマクラの集団が形成されたことが示された。このことから、現存のヒラノマクラは、かつて相模湾から野間岬沖を経由し、南西諸島海溝に到達したという分散過程を経た可能性がある。しかし、この順番での分散は黒潮の流れとは逆向きとなり、ヒラノマクラの幼生が黒潮の流れに逆らって分散していくことになる。

説明文の12
これには2つ理由が考えられます。 1つ目の理由として、解析に用いた相模湾のヒラノマクラが少なかったためかもしれません。 ●これは野間岬沖、南西諸島海溝、相模湾間の遺伝的多様性です。 ●相模湾の遺伝的多様性が、他2地点と比べて比較的低い値となりましたが、 ●これは、解析に用いた個体数が少なく、本研究で発見されなかったハプロタイプが相当数残っているためかもしれません。 このように相模湾のデータには他2地点のデータと比べると正確性に欠ける部分があるのかもしれません。
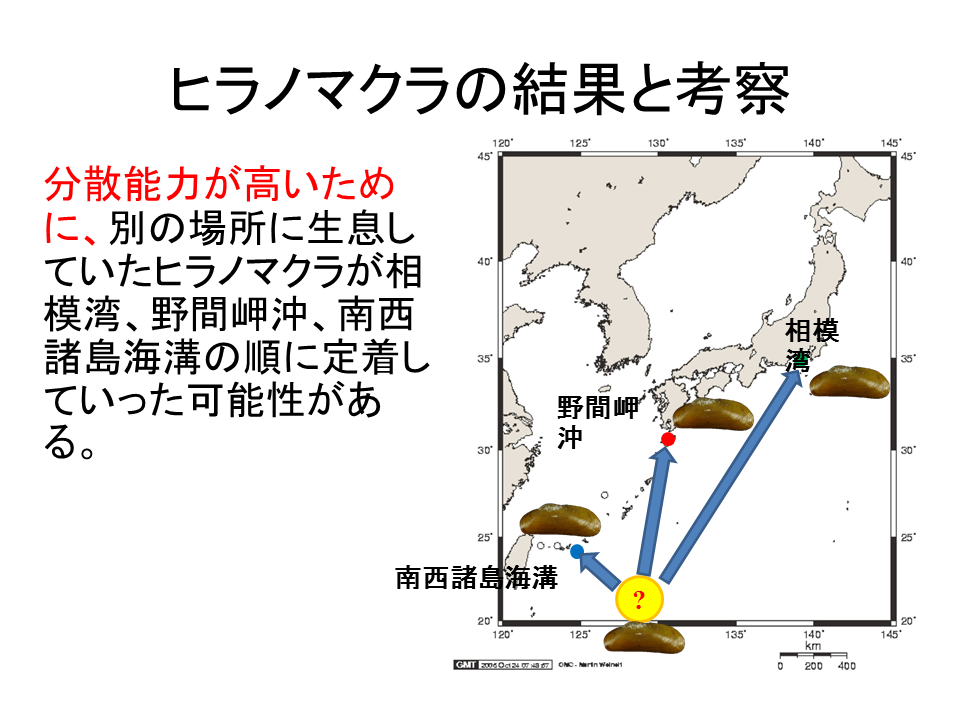
説明文の13
もう一つの可能性として、分散能力が高いために、別の場所に生息していたヒラノマクラが、相模湾、野間岬沖、南西諸島海溝の順に定着していったということが考えられる。
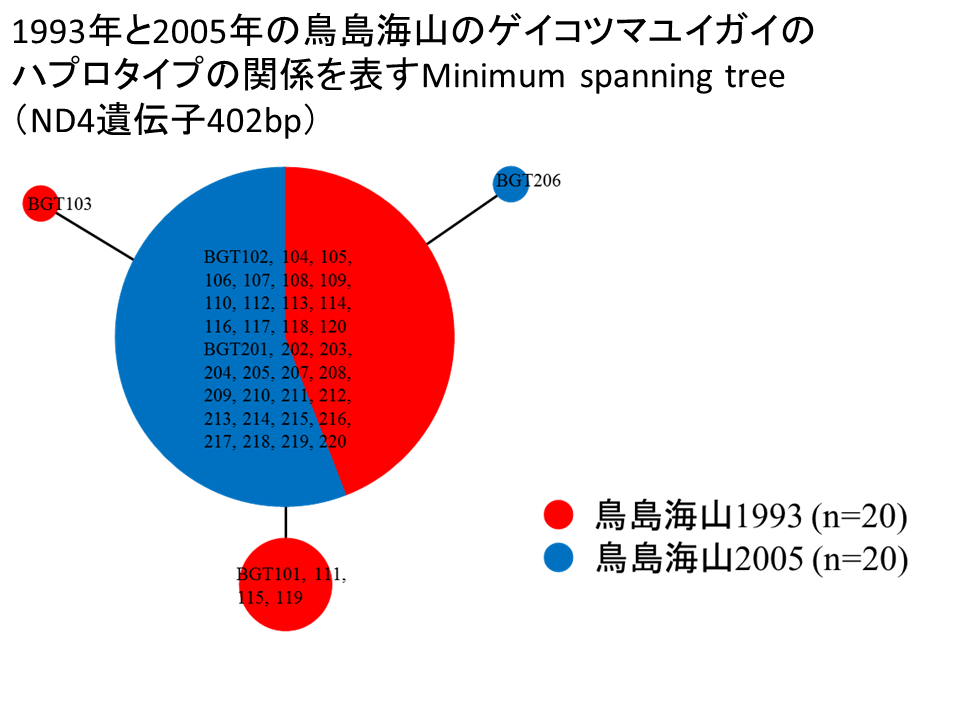
説明文の14
残念ながら、ゲイコツマユイガイは1か所でしか採取できなかったため、地点間での比較を行うことができない。経年変化について、1993年から2005年の鳥島海山のゲイコツマユイガイをそれぞれの年度でグループとして、ND4遺伝子402bpに基づくミニマムスパニングツリーを構築した。1993年を赤色、2005年を青色で示す。1993年と2005年の多くの個体が同じハプロタイプを共有した。また、12年間の時間の経過があるものの、ハプロタイプの数は4つしかなく、7年間の調査で26個のハプロタイプが見つかった野間岬沖のヒラノマクラと比べ、非常に少ない。

説明文の15
1993年と2005年のFst、Fst P value及び遺伝的多様性を算出した。Fstと遺伝的多様性は、1993年と2005年で有意差はなかった。

説明文の16
鳥島海山のゲイコツマユイガイは12年間で遺伝的な違いはほとんど見られなかった。このことから、安定した規模の大きな未知の集団から、同じ遺伝的組成をもつ個体が繰り返し移入してきた可能性がある。また、採取地点の周囲に他の生息場所が全くなかったことから、鳥島海山のゲイコツマユイガイは遠く離れたところから移入してきていると考えられ、ゲイコツマユイガイの分散能力が高いことが示唆される。
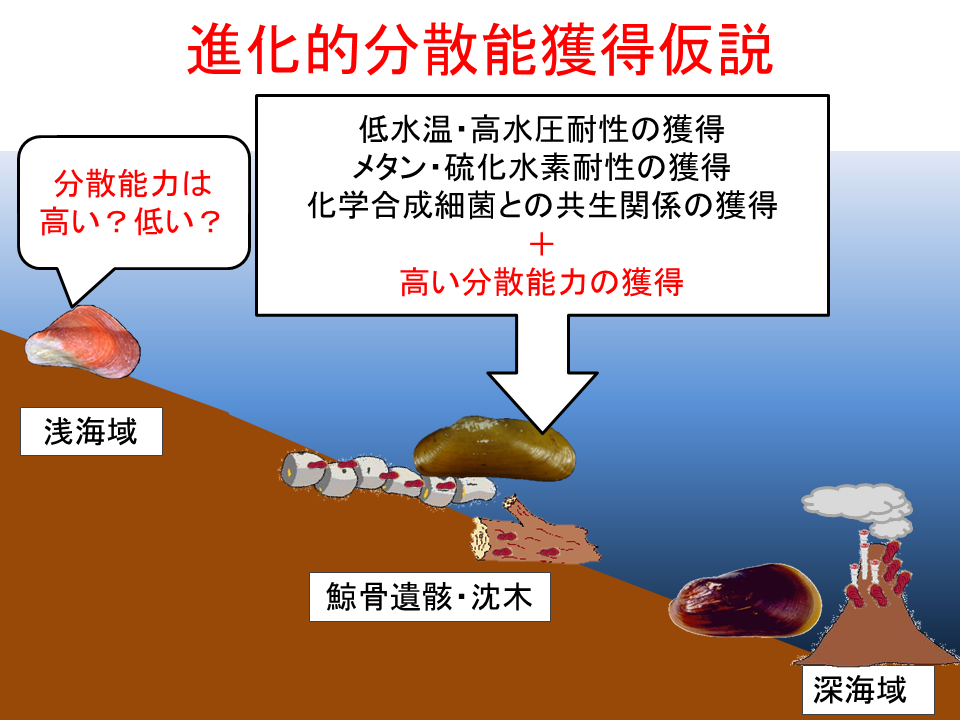
説明文の17
こ進化的分散能獲得仮説は、低水温・高水圧耐性の獲得、メタン・硫化水素耐性の獲得、化学合成細菌との共生関係の獲得と同じように、高い分散能力も鯨骨遺骸上で獲得したとする仮説である。この仮説を証明するためには、浅海に生息するイガイ類の分散能力を調べる必要がある。意外なことに、浅海のイガイ類の分散能力についての報告はほとんどない。